
Themen • Kurzbeiträge • Streiflichter
Vögel aus der Kreide – Busch statt Stammbaum
von Reinhard Junker
Studium Integrale Journal
28. Jahrgang / Heft 2 - September 2021
Seite 79 - 86
Zusammenfassung: Ein aktuelles Cladogramm* von Vogelgattungen aus der geologischen Periode der Kreide zeigt, dass ein Großteil der Fossilformen in einem relativ engen Zeitfenster (erstmals oder überhaupt) überliefert ist. Eine große Formenvielfalt erscheint fossil plötzlich im Aptium*. Die vergleichsweise wenigen geologisch älteren Gattungen aus dem Barremium und Hauterivium sind kaum als Vorläuferformen geeignet, im Gegenteil: Gerade einige der geologisch ältesten Gattungen sind „abgeleitet“ („höherentwickelt“) und stehen ziemlich tief eingeschachtelt (also weit oben) im Cladogramm. Der Gesamtbefund der in der Kreide überlieferten Vögel widerspricht deutlich evolutionstheoretischen Erwartungen.
• • • •
Mit einem Stern* versehene Begriffe werden im Glossar erklärt.
Abb. 1: Rekonstruktion von Archaeornithura. (Zongda Zhang / Institute of Vertebrate Paleontology and Paleoanthropology, Bejing) Einleitung
Die evolutive Ableitung der Vögel von Dinosauriern hat in der Fachwelt einen Status der Gewissheit ähnlich wie Evolution insgesamt als Rahmenparadigma. Im Jahr 1998 titelte Henry Gee in der Wissenschaftszeitschrift Nature dazu: „Birds and dinosaurs – the debate is over“ (Gee 1998). Widerspruch in der Fachwelt gibt es kaum (Feduccia 2020).
Kompakt
Die evolutive Ableitung der Vögel von Dinosauriern gilt in der Fachwelt als gesichert. Dennoch liegen systematische Befunde vor, die einer evolutionstheoretischen Deutung der relevanten Fossilfunde entgegenstehen. Einer dieser Befunde ist das ziemlich plötzliche Auftreten verschiedenster Vogelgattungen in großer Formenfülle, was einer schrittweisen Evolution widerspricht. Zudem zeigt sich, dass die geologisch ältesten Formen, die zu den beiden Hauptgruppen der kretazischen* Vögel gehören, in der Regel nicht wie evolutionstheoretisch zu erwarten an der Basis, sondern an unterschiedlichen abgeleiteten Positionen im hypothetischen Stammbaum positioniert sind.
In einer Reihe von Übersichtsartikeln des letzten Jahrzehnts wird darüber hinaus behauptet, dass auch nachvollziehbar sei, dass die typischen Vogelmerkmale schrittweise in Dinosaurier-Vorläufer-Gruppen entstanden seien. Eine genauere Analyse zeigt jedoch, dass die Verhältnisse sehr viel komplizierter sind: Zahlreiche Vogelmerkmale kommen zwar in unterschiedlichen Dinosaurier-Gruppen der Theropoden vor, aber es muss in vielen Fällen von einer konvergenten und damit von den Vögeln unabhängigen Entstehung dieser Merkmale ausgegangen werden (Junker 2019), meist sogar mehrfach, und das oft in Gattungen, die deutlich jünger sind als die ältesten Vogelgattungen. Mit anderen Worten: Die Vogelmerkmale bei Dinosauriern passen in vielen Fällen weder im Cladogramm* noch stratigraphisch* als Vorläufermerkmale (Abb. 2).
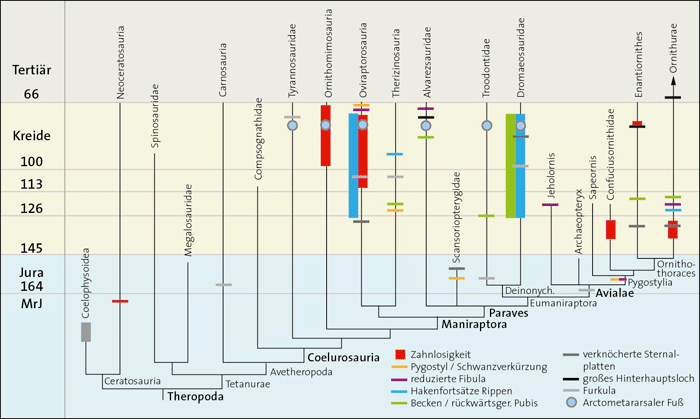
Abb. 2: Zeitskaliertes Cladogramm der Theropoden-Dinosaurier-Gruppen und Vögel und Zusammenschau der konvergenten Verteilung von neun vogeltypischen Merkmalen. Die stammesgeschichtlichen Rekonstruktionen verschiedener Bearbeiter ergeben häufig unterschiedliche Anordnungen einzelner Gruppen. An der vielfachen Konvergenz ändern die verschiedenen Cladogramm-Varianten jedoch nichts Wesentliches. Die Merkmalsverteilungen sind vielfach so unsystematisch, dass die Annahme zahlreicher Konvergenzen unvermeidbar ist. MrJ: Millionen radiometrische Jahre. (Nach zahlreichen Quellen zusammengestellt, siehe dazu Junker 2019)
Feduccia (2020) vertritt die Auffassung, dass es sich bei zahlreichen Dinosaurier-Gattungen mit Vogelmerkmalen um sekundär flugunfähig gewordene Formen handelt; eine Hypothese, die gelegentlich auch von anderen Wissenschaftlern diskutiert wurde (z. B. Paul 2002). Trifft dies zu, würden cladistische Analysen (Stammbaumrekonstruktionen) in die Irre führen, weil durch die cladistische Methode Rückentwicklungen oft nicht erkannt werden können.
Vogelfossilien sind ab dem Oberjura bekannt, allen voran der berühmte „Urvogel“ Archaeopteryx. In noch älteren Schichten des Oberjura wurde der vierflügelige Anchiornis entdeckt, der wie Archaeopteryx voll befiedert war und dessen Art des Flugs ungeklärt ist (Hu et al. 2009; Wang et al. 2017b). Auch die Gattungen Caihong (Hu et al. 2018) und Serikornis (Lefèvre et al. 2017) sind hier zu nennen. Gewöhnlich werden Anchiornis und andere befiederte Gattungen aus dem Oberjura als „befiederte Dinosaurier“ interpretiert, doch das folgt nicht aus den Befunden, sondern aus der theoretischen Vorstellung einer Abstammung der Vögel von Dinosauriern. Darauf soll an dieser Stelle nicht weiter eingegangen werden (siehe Diskussion bei Junker 2017).
Während im Oberjura voll befiederte Gattungen eher dünn gesät sind, sind in der Unterkreide vor allem ab dem Aptium (ca.126–113 Millionen radiometrische Jahre [MrJ]) sehr viele Vogelgattungen fossil überliefert. In einer aktuellen Überblicksarbeit haben Wang et al. (2021) ein Cladogramm (Ähnlichkeitsbaum) der im Jura und in der Kreide fossil überlieferten Vogelgattungen veröffentlicht. In diesem Cladogramm ist auch angegeben, in welchen Schichten die Fossilfunde überliefert sind (Abb. 3). Schon ein flüchtiger Blick zeigt, dass ein Großteil der kretazischen* Gattungen in der Unterkreide nur im Aptium fossil vertreten ist und dass sehr viele Gattungen annähernd gleichzeitig auftreten. Das gilt insbesondere für Vertreter der beiden Großgruppen der Enantiornithes* (Gegenvögel) und Ornithuromorpha* („Vogelschwänze“). Darauf weisen auch Wang et al. (2021,1) hin: Bereits „kurz nach der Abspaltung von anderen Theropoden im späten Jura diversifizierten die Vögel des Mesozoikums in zwei Hauptgruppen – die Enantiornithes und die Ornithuromorpha“. Ihre Ergebnisse zeigen, dass diese beiden Hauptgruppen der Vögel des Mesozoikums in einem diskreten Merkmalsraum unterscheidbar sind. Beide Gruppen werden als Ornithothoraces* („Brustbein-Vögel“) zusammengefasst.
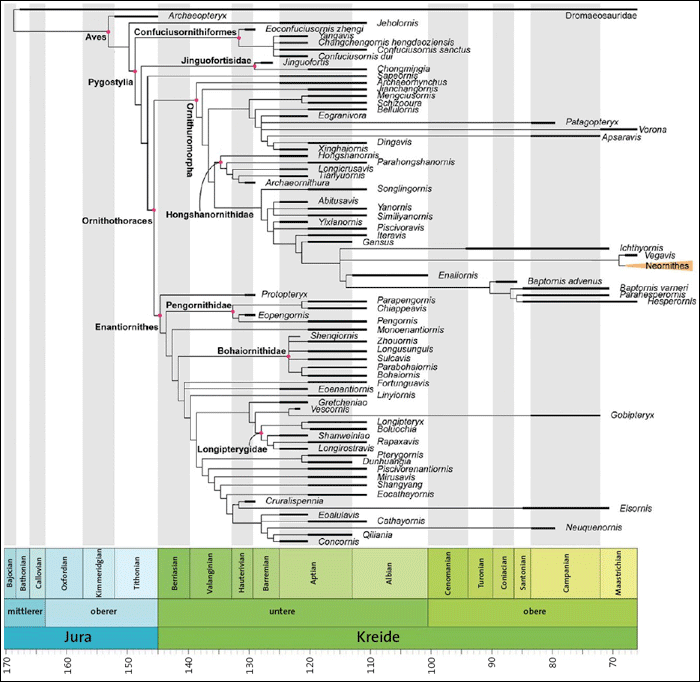
Abb. 3: Cladogramm der in der geologischen Formation der Kreide überlieferten fossilen Vogelgattungen mit Darstellung (Balken), in welchen Stufen sie gefunden wurden. Die Zahlen am unteren Ende geben Millionen radiometrische Jahre an. (Nach Wang et al. 2021; mit freundlicher Genehmigung)
Nur wenige Vogelgattungen sind in älteren Schichten des Hauteriviums und des Barremiums gefunden worden (Abb. 3). Evolutionstheoretisch würde man vermuten, dass diese Formen Merkmalskonstellationen aufweisen, die eine Interpretation als Vorläuferformen erlauben. Im Folgenden sollen die betreffenden Gattungen daraufhin untersucht werden.
Glossar
Alula: Daumenfittich; Federn am Daumen des Vogelflügels, die besonders beim Bremsen und beim Landeanflug wichtig sind. Aptium: Stufe der Unterkreide, auf ca. 126-113 → MrJ datiert. Cladogramm: Ähnlichkeitsbaum, der meist als Abstammungsbaum interpretiert wird. Enantiornithes: Gegenvögel, eine der beiden großen formenreichen Vogelgruppen, die in der geologischen Formation der Kreide fossil überliefert sind. Es handelt sich meist um baumlebende Arten. Ihre Bezeichnung rührt daher, dass bei ihnen die Gelenke zwischen dem Schulterblatt und dem Rabenbein im Vergleich mit der anderen in der Kreide vorkommenden Vogelgruppe, den → Ornithuromorphen (zu denen auch die heute lebenden Vögel gerechnet werden), genau anders herum angeordnet sind: Bei den Gegenvögeln liegt am Schulterblatt eine Gelenkpfanne und am Rabenbein ein Gelenkzapfen vor; bei den Ornithuromorphen ist es genau umgekehrt. Diese beiden gegensätzlichen Konstellationen können kaum evolutionär auseinander hervorgegangen sein; deshalb werden die beiden Gruppen in zwei getrennte Linien gestellt. Geisterlinie: Von Geisterlinien spricht man, wenn ein → Cladogramm gemäß evolutionstheoretischer Deutung die Annahme längerer Zeiträume erfordert, in denen die betreffenden Linien existiert haben müssen, ohne dass sie fossil belegt sind; in Abb. 3 z. B. sehr ausgeprägt bei Patagopteryx, Vorona und Apsaravis. Humerus: Oberarmknochen. kretazisch: zur geologischen Formation der Kreide gehörend. MrJ: Millionen radiometrische Jahre; Jahre gemäß radiometrischen Altersbestimmungen. Ornithothoraces: „Brustbein-Vögel“. Vogelgruppe aus → Enantiornithes und → Ornithuromorpha. Der Name Ornithothoraces bezieht sich auf das Vorhandensein eines gut entwickelten Brustbeinkiels, der eine größere Oberfläche zur Befestigung der Flugmuskulatur ermöglicht. Ornithuromorpha: „Vogelschwänze“; für diese Gruppe gibt es keine eingebürgerte deutsche Bezeichnung, der Name bezieht sich auf den für heutige Vögel typischen Fächerschwanz. Die Ornithuromorpha bilden neben den → Enantiornithes die zweite in der Kreide überlieferte Vogelgruppe; die Arten lebten meist in Wassernähe. plesiomorph: evolutionstheoretische Deutung eines Merkmals als ursprünglich. Pygostyl: miteinander verwachsene letzte Schwanzwirbel der Vögel; daran sitzen die Schwanzfedern. stratigraphisch: geologische Schichten betreffend.
Die geologisch ältesten kretazischen Vogelgattungen
Archaeornithura. Diese zu den Ornithuromorpha gehörende amselgroße Gattung aus der Jehol-Gruppe Nordchinas ist der älteste fossile Nachweis dieser Vogelgruppe (Alter auf 130,7 MrJ bestimmt). Dennoch erweist sie sich als überraschend „modern“ (Wang et al. 2015; Abb. 1, Seite 11). Archaeornithura besaß ein ziemlich „modernes“ Federkleid mit asymmetrischen Schwungfedern und gut entwickelter Alula* (Daumenfittich) mit mindestens drei Federn und fächerförmigen Schwanzfedern, die denjenigen heutiger Vögel erstaunlich ähnlich sind (Wang et al. 2015, 7). Beide Merkmale sind bei heutigen Vögeln für langsamen Flug und Manövrierbarkeit aerodynamisch wichtig. Archaeornithura dürfte daher ein guter Flieger gewesen sein. Die Knochen am Ende der Flügel sind stark verschmolzen und es ist eine U-förmige Furkula (verwachsenes Schlüsselbein) ausgebildet (Balter 2015) – beides Kennzeichen auch heutiger Vögel. Seine langen Beine waren nicht befiedert, was darauf hinweist, dass es sich um einen Watvogel gehandelt haben dürfte, der im flachen Wasser herum stakste und dort nach Nahrung suchte.
Die Position in der Schichtenabfolge und die Position im evolutionstheoretischen Stammbaum passen oft nicht zusammen.
Archaeornithura ist im Cladogramm der Gruppe tief eingeschachtelt (also weit oben; Wang et al. 2015, 6), also evolutionstheoretisch „hochentwickelt“ („abgeleitet“); die neue Gattung ist innerhalb der Hongshanornithidae eine spezialisierte Form, wobei diese Familie ihrerseits innerhalb der Ornithuromorpha ebenfalls tief eingeschachtelt ist, anders als andere Taxa aus jüngeren Schichten (also entgegen der zu erwartenden Reihenfolge). Wang et al. (2015, 6) sprechen von Inkonsistenzen zwischen Stratigraphie und Phylogenie, was die Annahme sogenannter Geisterlinien* erforderlich mache.

Abb. 4: Protopteryx-Fossil aus Fengning, Hebei, China (Tiouraren, CC BY-SA 4.0) und Rekonstruktion (es.dino.wikia.com, CC-BY-SA).
Protopteryx. Protopteryx (Abb. 4) ist eines der ältesten Fossilien der Gegenvögel und wird auf 130–129 MrJ datiert (O’Connor et al. 2020). Die Gattung nimmt zwar im Cladogramm eine basale Position ein, weist aber auch eine Reihe von Merkmalen auf, die für eine sehr gute Flugfähigkeit sprechen – trotz deutlich verschiedener Anatomie im Vergleich mit heutigen Vögeln. Navalón et al. (2015) stellen fest, dass Protopteryx fengningensis Vorderextremitäten mit „modernen“ Proportionen besaß, außerdem ein gekieltes Brustbein, einen „fortschrittlichen“ Schultergürtel mit einem Kanal für die Passage der Flugmuskeln (Dreiknochenkanal, triosseal canal), was alles auf die Fähigkeit zu aktivem Flug und Flügelschlag ähnlich dem heutiger Vögel hinweise. Auch Ähnlichkeiten der Körperbedeckung mit heutigen Vögeln mit identischer Federanordnung würden in diese Richtung weisen. Chiappe et al. (2019) fanden an einem gut erhaltenen Exemplar Merkmale, die auf die Fähigkeit zu stoßweisem Flug („bounding“ oder „flap-gliding“) hindeuten, was das früheste Auftreten einer solchen energiesparenden Flugstrategie markiere, wie es auch bei heutigen Vögeln bekannt ist.
Protopteryx weist Merkmale auf, die sonst bei den Gegenvögeln fehlen, jedoch bei den Ornithuromorpha verbreitet sind, wie z. B. das Procoracoid (Teil des Schultergürtels) und die seitlichen Fortsätze am Rabenbein und der körpernah konvexe Kopf des Oberarmknochens (Humerus).
Der Humerus bei Protopteryx kann nach O’Connor et al. (2020, 15) als plesiomorphe* (ursprüngliche) Ausprägung der Ornithothoraces* gewertet werden, ebenso der kleine Procoracoid-Fortsatz und ein seitlicher Fortsatz am Humerus. Diese Merkmale könnten aber auch unabhängig voneinander in der Protopteryx-Linie und den Ornithuromorphen entstanden sein, da ein Procoracoid-Fortsatz auch unabhängig in der von beiden Gruppen unabhängigen Jeholornis-Linie ausgebildet ist und ein seitlicher Fortsatz am Humerus im basalen Ornithuromorphen Schizooura fehlt.

Abb. 5: Eopengornis-Fossil mit langen Schwanzfedern. (Aus Wang et al. 2014; CC BY-SA 4.0)
O’Connor et al. (2020, 2) weisen darauf hin, dass die Vogelwelt aus dem Protopteryx-Horizont morphologisch recht vielfältig ist, obwohl es sich um das früheste fossil dokumentierte Stadium der Gegenvögel-Evolution handelt. Das spreche dafür, dass diese Vogelgruppe zeitlich früher entstanden ist. Es seien zahlreiche morphologische Merkmale vorhanden, die bei jüngeren Gegenvögeln fehlen, aber charakteristisch für Ornithuromorphen sind: ein pflugförmiges Pygostyl* von Cruralispennia (s. u.), das dem Pygostyl von Sapeornis ähnelt, ein Procoracoid-Fortsatz bei Protopteryx und ein kugelförmiger Humeruskopf* von Eopengornis (s. u.) und anderen Pengornithiden. Der Umstand, dass diese Vogelgruppen durch häufiges Vorkommen von Homoplasien charakterisiert sind, „erschwert die Versuche, ihre Phylogenie zu verstehen, wobei aktuelle phylogenetische Analysen nur schwach unterstützt werden und instabil sind“ (O’Connor et al. 2020, 15).
Eopengornis. Diese Gattung gehört ebenfalls zu den ältesten Vögeln aus der Kreide (Jehol-Gruppe China, ca. 131 MrJ) (Wang et al. 2014). Die Form des Schwanzes ist ungewöhnlich: Ein Paar verlängerter, vollständig gefiederter, von der zentralen Achse (Rachis) dominierter Federn (Sullivan et al. 2016; vgl. Abb. 5). Das stütze nach Wang et al. (2014, 805) die Hypothese, dass Rachis-dominierte Federn („racket-plumes“) in basalen Vögeln modifizierte Konturfedern darstellen und nicht verlängerte Schuppen (Wang et al. 2014, 805). Aufgrund morphologischer Ähnlichkeiten mit basalen Ornithuromorphen und aufgrund der Position im Cladogramm wird die Gattung als „primitiv“ eingestuft. Wang et al. (2020, 811f.) nennen „beispielsweise einen konvexen vorderen Rand und eine flache kraniale Oberfläche des Humerus, ein kurzes Hypocleidium und den Bau des Brustbeins“. Abgesehen von den Schwanzfedern, die vermutlich als Schmuckfedern fungierten, scheint diese Gattung also in eine basale Stellung zu passen.
Cruralispennia. Die zu den Gegenvögeln gestellte Art Cruralispennia multidonta (Abb. 6) besitzt Merkmale, die typisch sind für die Ornithuromorpha, weist also einen Mix aus Merkmalen beider Vogelgruppen auf. Die Gattung erhielt ihren Namen aufgrund der ungewöhnlich befiederten Beine (crus = Schenkel, pennae = Federn). Sie gehört nach Archaeopteryx zu den ältesten Vögeln (Huajiying-Formation Nordwestchinas, 131 MrJ), nimmt aber dennoch eine abgeleitete Stellung unter den Gegenvögeln ein und kann daher nicht als Übergangsform interpretiert werden. Nach phylogenetischen Analysen steht die Gattung tief eingeschachtelt weit oben im Cladogramm, was „unerwartet“ ist (Wang et al. 2017a, 1), weil evolutionär bei den ältesten Formen natürlich mit „primitiven“ Merkmalen gerechnet wird. Wang et al. (2017a) sprechen von einer „stratigraphisch-phylogenetischen Diskrepanz“.

Abb. 6: Rekonstruktion von Cruralispennia. (Fanboyphilosopher, CC BY-SA 4.0)
Besonders überraschend ist der Besitz eines pflugförmigen Pygostyls, dessen Form für Ornithuromorpha typisch ist (Gegenvögel haben sonst ein längliches Pygostyl). Wang et al. (2017a) nehmen an, dass es konvergent zu den Ornithuromorpha entstanden ist. Ebenfalls bisher nicht bekannt unter den Gegenvögeln der Unterkreide war die Ausbildung eines extrem schlanken Rabenbeins (Coracoid, Element des Schultergürtels); bisher war diese Form des Rabenbeins nur bei Formen aus der Oberkreide bekannt. Ungewöhnlich sind auch die bereits erwähnten Federn an den Schenkeln, die im körpernahen Bereich drahtartig sind und distal fädige Spitzen besitzen. Diese Federform war bisher unbekannt und erweitert das ohnehin bereits reichhaltige Spektrum von Federformen bei frühen Vögeln, das die Vielfalt heutiger Federtypen noch übersteigt. Histologische Befunde an den Knochen deuten darauf hin, dass Cruralispennia schnell wuchs und vermutlich binnen eines Jahres ausgewachsen war; für die Gegenvögel ein weiteres ungewöhnliches, als „abgeleitet“ interpretiertes Merkmal.
Der neue Fund trägt zum Bild eines plötzlichen fossilen Erscheinens einer erheblichen Formenvielfalt in der Unterkreide bei.

Abb. 7: Rekonstruktion von Jinguofortis. (© Chung-Tat Cheung)
Jinguofortis. Diese etwa krähengroße Gattung aus Hebei (Nordost-China, auf 127 MrJ datiert, Abb. 7) besaß zwar wie heutige Vögel ein Pygostyl (verschmolzene Schwanzwirbel), wird aber dennoch mit unklarer systematischer Position außerhalb der Ornithothoraces in eine basale Position unter den Vögeln der Kreide gestellt (Wang et al. 2016). Ein weiteres ausgesprochen „modernes“ Merkmal sind die stark reduzierten Finger. Neben vogeltypischen Merkmalen kommen aber auch Merkmale vor, die sonst bei Theropoden-Dinosauriern bekannt und vogeluntypisch sind. Dazu gehören Krallen an den Fingern der Flügel, eine bumerangförmige, vermutlich starre Furkula (Gabelbein) anstelle einer flexiblen Ausprägung, ein Kiefer mit winzigen Zähnen statt eines Schnabels und ein verschmolzener Schultergürtel (Schulterblatt und Rabenbein). Da letzteres Merkmal ungünstig für das Fliegen erscheint, weil es die Flexibilität für den Schlagflug einschränkt, erhielt die Art ihren Artnamen „perplexus“. Nichtsdestotrotz waren die breiten, kurzen Flügel von Jinguofortis typisch für Vögel, die gut zwischen Bäumen manövrieren können. Vielleicht war eine bisher unbekannte Art des Fliegens verwirklicht. Bei heutigen Vögeln ist eine vergleichbare Fusion der beiden Schultergelenksknochen nur bei flugunfähigen Formen bekannt. Dass auch Jinguofortis sekundär flugunfähig war, scheint jedoch eher unwahrscheinlich, zumal die Schwungfedern ausgeprägt asymmetrisch waren (Wang et al. 2018, 10710).
Der verschmolzene Schultergürtel (Schulterblatt und Rabenbein) ähnelt zwar der Situation bei einigen Theropoden-Dinosauriern, trotzdem eignet sich dieses Merkmal nicht als Beleg für eine stammesgeschichtliche Verbindung von Dinosauriern und Vögeln, weil bei Archaeopteryx diese beiden Knochen nicht verschmolzen waren. Daher wird eine unabhängige (konvergente) Entstehung angenommen, und zudem ebenfalls konvergent bei Confuciusornis (Wang et al. 2016; 2018).
Der Besitz von Zähnen ist bei mesozoischen Vögeln verbreitet und auch bezüglich dieses Merkmals ist die Ausprägung bei Jinguofortis evolutionstheoretisch nicht gut passend. Denn bei Jinguofortis waren Prämaxilla (Zwischenkiefer) und Maxilla (Oberkieferknochen) bezahnt, während bei Archaeopteryx und den Ornithothoraces die Spitze der Prämaxilla und bei Jeholornis die ganze Prämaxilla zahnlos ist. Daher müsste bezüglich dieses Merkmals evolutionstheoretisch eine Rückentwicklung bzw. wie beim Schultergürtel eine Art evolutionärer Zickzackkurs angenommen werden, was allgemein als unplausibel gilt. Die Merkmalsverteilung passt somit nicht in ein hierarchisches eingeschachteltes System; daher müssen auch hier Konvergenzen angenommen werden. Das Merkmalsmosaik bei Jinguofortis ist so gestaltet, dass die Gattung insgesamt nicht als evolutionäre Übergangsform passt, sondern in eine eigene evolutive Linie gestellt werden muss.
Orienantius. Diese Gattung fehlt zwar im Cladogramm von Wang et al. (2021), verdient hier aber auch Erwähnung, weil sie ebenfalls zu den ältesten Gegenvögeln gerechnet wird (Huajiying Formation; Hauterivium, ca. 131 MrJ). Auch Orienantius besitzt Merkmale, die für abgeleitete Gegenvögel charakteristisch sind – entgegen der Altersstellung (Liu et al. 2019). Bemerkenswert ist die Erhaltung von Flughäuten, was darauf hindeutet, dass diese Vögel einen sehr gut manövrierfähigen, stoßweisen Flatter-Gleitflug ausüben konnten.
Orienantius ritteri teilt mehrere Merkmale mit jüngeren und abgeleiteten Gegenvögeln einschließlich einem spezialisierten Brustbein, einem langen und schmalen Pygostyl, einer geringen Anzahl von Zähnen im Oberkiefer und einem kurzen Alula-Finger. Die erhaltene Kontur der Beinmuskulatur ist mit der von modernen Vögeln identisch, und die Weichteile um die Fußknochen deuten darauf hin, dass die Füße von Orienantius ritteri keine größeren Muskeln besaßen, was ebenfalls der Situation bei heutigen Vögeln entspricht (Liu et al. 2019, 204).
Liu et al. (2019, 206) stellen zusammenfassend fest, dass Orienantius zahlreiche Merkmale aufweist, die für spätere Taxa typisch sind. Die Gattung unterstreiche die Vielfalt, die die Gegenvögel zu Beginn ihrer Fossilüberlieferung aufwiesen (Liu et al. 2019, 191).
Weitere Vogelgattungen, die nicht zu den Ornithothoraces gehören. Einige kretazische Gattungen werden evolutionstheoretisch als primitiver eingestuft als die zu den Ornithothoraces zusammengefassten Gegenvögel und Ornithuromorpha. Dazu gehören Confuciusornis, Eoconfuciusornis, Jeholornis und Sapeornis. Diese Gattungen sind mit Ausnahme von Eoconfuciusornis wie die meisten anderen kretazischen Vögel erst ab dem Aptium fossil überliefert, so dass eine Unstimmigkeit von Phylogenie und Stratigraphie außer bei Eoconfuciusornis auch bei diesen Gattungen gegeben ist.
Schlussfolgerungen
Nur eine der sechs hier diskutierten ältesten kretazischen Vogel-Gattungen kann aufgrund ihrer Merkmalsausprägungen relativ widerspruchsarm in eine basale Position der Ornithothoraces gestellt werden; mit einigen Abstrichen gilt das auch für Protopteryx. Die anderen Gattungen passen eindeutig nicht ins evolutionstheoretische Bild, wonach die ältesten Formen auch die evolutionär ursprünglichsten (plesiomorphen) Formen sein sollten. Vielmehr werden vier der sechs betreffenden Gattungen als evolutionär abgeleitet interpretiert. Die stratigraphischen Positionen und die aufgrund der jeweiligen Merkmalskonstellationen ermittelten phylogenetischen Stellungen passen bei Weitem nicht zusammen. Wie erwähnt kommentieren die Forscher diese Befunde als Inkonsistenzen zwischen Stratigraphie und Phylogenie (Wang et al. 2015, 6, bei Archaeornithura), als „stratigraphisch-phylogenetische Diskrepanz“ (Wang et al. 2017a, bei Cruralispennia) oder merken an, dass aktuelle phylogenetische Analysen instabil seien (O’Connor et al. 2020, 15).
Die meisten älteren Gattungen passen nicht ins Bild, wonach die ältesten Formen auch die evolutionär ursprünglichsten Formen sein sollten.
Zusammen mit dem Befund, dass ab dem Aptium plötzlich eine erhebliche Vielfalt von Vogelgattungen fossil überliefert ist (Abb. 3), passt der Gesamtbefund der stratigraphischen Abfolgen der Vogelgattungen der Kreide nicht zu einer evolutionstheoretischen Deutung der Vogelfossilien der Kreide. Dass es – unter der Annahme von Makroevolution – Lücken in der Fossilüberlieferung gibt, ist nachvollziehbar. Die Lücken scheinen hier aber systematischer Natur zu sein, so dass in großem Umfang Geisterlinien angenommen werden müssen. Daher ist aufgrund der fossilen Daten eine kritische Betrachtung des zugrundeliegenden Evolutionsmodells angebracht. Dies umso mehr, als viele Theropoden-Dinosaurier, die als Vogelvorläufer gewertet werden oder die vogeltypische Merkmale aufweisen, auch nicht geologisch älter und in vielen Fällen sogar geologisch jünger sind als ein Großteil der Vögel aus der Kreide (Junker 2017).
Aus der Sicht der Schöpfungslehre: Warum sind in der Kreide andere Vogeltypen fossil überliefert als im Tertiär? Die Enantiornithes sind nur in kretazischen Schichten überliefert. Die Ornithuromorpha überlebten zwar die Kreide-Tertiär-Grenze, aber die tertiären Vogelgruppen unterscheiden sich deutlich von den Ornithuromorpha der Kreide. Wie kann dieser Befund erklärt werden, wenn man gemäß der biblischen Schöpfungslehre von einer in Jahrtausenden bemessenen Erdgeschichte ausgeht und von einer Erschaffung aller Grundtypen in der Schöpfungswoche? Warum ist die Verteilung der Vogelgruppen trotz gleichzeitiger Existenz der Grundtypen in Kreide und Tertiär bei jeweils weltweiter Verbreitung der beiden Großgruppen so verschieden? Warum ist die Vogelwelt in den Ablagerungen des Jura und in der Kreide eine ganz andere als die des Tertiärs? Die Frage muss als ungelöstes Problem der biblischen Schöpfungslehre konstatiert werden. Beide Ursprungsmodelle – Evolution und biblische Schöpfungslehre – haben mit zwar sehr verschiedenen, aber gleichermaßen schwerwiegenden Problemen zu kämpfen.
When modern birds took flight. Science 348, 617.
Anatomy and flight performance of the early enantiornithine bird Protopteryx fengningensis: Information from new specimens of the Early Cretaceous Huajiying formation of China. Anat. Rec. 303, 716–731, doi:10.1002/ar.24322.
Romancing the birds and dinosaurs. Irvine, Boca Raton.
Birds and dinosaurs – the debate is over. Nature, doi:10.1038/news980702-8.
A pre-Archaeopteryx troodontid theropod from China with long feathers on the metatarsus. Nature 461, 460-463.
A bony-crested Jurassic dinosaur with evidence of iridescent plumage highlights complexity in early paravian evolution. Nat. Comm. 9:217.
Dino-Federvieh – Zum Ursprung von Vogelfeder und Vogelflug. https://www.wort-und-wissen.org/wp-content/uploads/b-17-1_feder-und-flug.pdf.
Sind Vögel Dinosaurier? Eine kritische Analyse fossiler Befunde. https://www.wort-und-wissen.org/wp-content/uploads/b-19-4_dinos-voegel.pdf.
A new Jurassic theropod from China documents a transitional step in the macrostructure of feathers. Sci. Nat. 104:74.
Soft tissue preservation in two new enantiornithine specimens (Aves) from the Lower Cretaceous Huajiying Formation of Hebei Province, China. Cretaceous Res. 95, 191–207, doi:10.1016/j.cretres.2018.10.017.
Soft-tissue and dermal arrangement in the wing of an Early Cretaceous bird: Implications for the evolution of avian flight. Sci. Rep. 5:14864.
New information on the plumage of Protopteryx (Aves: Enantiornithes) from a new specimen. Cretaceous Res. 116, 104577.
Dinosaurs of the air. The Evolution and Loss of Flight in Dinosaurs and Birds. Johns Hopkins University Press.
Complexities and novelties in the early evolution of avian flight, as seen in the Mesozoic Yanliao and Jehol Biotas of Northeast China. Palaeoworld, http://dx.doi.org/10.1016/j.palwor.2016.12.001.
Insights into the evolution of rachis dominated tail feathers from a new basal enantiornithine (Aves: Ornithothoraces). Biol. J. Linn. Soc. 113, 805-819.
The oldest record of ornithuromorpha from the early cretaceous of China. Nat. Comm. 6:6987, doi: 10.1038/ncomms7987.
A new basal bird from China with implications for morphological diversity in early birds. Sci Rep. 6:19700, doi: 10.1038/srep19700.
A bizarre Early Cretaceous enantiornithine bird with unique crural feathers and an ornithuromorph plough-shaped pygostyle. Nat. Comm. 8:14141, doi: 10.1038/ncomms14141.
Basal paravian functional anatomy illuminated by high-detail body outline. Nat. Comm. 8:14576, doi: 10.1038/ncomms14576.
A new clade of basal Early Cretaceous pygostylian birds and developmental plasticity of the avian shoulder girdle. Proc. Natl. Acad. Sci. 115, 10708-10713.
The patterns and modes of the evolution of disparity in Mesozoic birds. Proc. R. Soc. B 288: 20203105; doi:10.1098/rspb.2020.3105.
Themen | Kurzbeiträge | Streiflichter
Studiengemeinschaft WORT und WISSEN e.V.
Letzte Änderung: 11/22/21
Webmaster