
Themen • Kurzbeiträge • Streiflichter
Vogelflug mehrfach entstanden?
von Reinhard Junker
Studium Integrale Journal
28. Jahrgang / Heft 2 - September 2021
Seite 107 - 109
Zusammenfassung: Nach einer aktualisierten Stammbaumrekonstruktion der Paraves (Vögel und deren nächsten Verwandten) muss eine mindestens dreimalige Entstehung des aktiven Flugs mittels Federkleid angenommen werden. Das widerspricht bisherigen evolutionstheoretischen Vorstellungen und spricht für eine freie Kombinierbarkeit von Merkmalen.
• • • • • •
Einführung
Abb. 1: Rekonstruktion des vierflügeligen Microraptor. (durbed.deviantart.com, CC BY-SA 3.0)
Eine mehrfach unabhängige (konvergente*) evolutive Entstehung der Flugfähigkeit mithilfe von Federn gehört zu den Dingen, die die meisten Biologen noch vor 20 oder 30 Jahren für ausgeschlossen erklärt hätten. Vogelfedern und Vogelflug sind dafür einfach zu speziell und zu komplex. Solche komplexen Merkmale und die mit ihnen verknüpften Fähigkeiten gelten als Schlüsselmerkmale, die als typisch für einen größeren Formenkreis angesehen werden. Die Herausforderung ist für Evolutionstheorien schon groß genug, die einmalige Entstehung von komplexen Merkmalen zu erklären. Die mehrfach unabhängige Entstehung – die evolutionstheoretisch immer ohne jede Zielorientierung beschrieben werden müsste – verschärft dieses Problem erheblich.
Mit einem Stern* versehene Begriffe werden im Glossar erklärt.
Dazu kommt die Schwierigkeit, Abstammungsverhältnisse zu rekonstruieren, wenn man sich nicht einmal auf Schlüsselmerkmale als Marker für gemeinsame Vorfahren verlassen kann. Schon 1998 beklagte Neil Shubin angesichts der mosaikartigen Merkmalsverteilung bei den geologisch ältesten Vierbeinern: „Wenn eine unabhängige Evolution von Schlüsselmerkmalen verbreitet ist, wie kann dann die Phylogenie rekonstruiert werden?“ (Shubin 1998, 13)1 Später stellte sich heraus, dass auch das für Säugetiere charakteristische Merkmal des Besitzes von drei Gehörknöchelchen – Hammer, Amboss und Steigbügel – unter fossilen Säugetieren mosaikartig verteilt ist, so dass mittlerweile auch bei diesem Schlüsselmerkmal evolutionstheoretisch eine mehrfach unabhängige Entstehung angenommen werden muss (Rich et al. 2005).
Nun muss auch der aktive Flug mithilfe von Federn in die Reihe der konvergenten Schlüsselmerkmale aufgenommen werden. Auch diese Fähigkeit soll nach einer neuen detaillierten Analyse mehrfach unabhängig evolutiv entstanden sein – genauer gesagt: mindestens dreimal. Es wird derzeit allerdings nicht angenommen, dass flächige, für den aktiven Flug taugliche Federn konvergent entstanden sind, sondern ausgehend von verschiedenen Formen mit flächigen Federn der aktive Schlagflug. Aber auch der Übergang von Formen mit Federn, die allenfalls zu einem Gleitflug befähigt waren, zu solchen, die aktiv fliegen konnten, erfordert erhebliche Umbauten und viele neue Fähigkeiten.

Abb. 2: Rekonstruktion von Rahonavis ostromi. (Nobu Tamura, CC BY 2.5) Welche Befunde führen zu den neuen Vorstellungen?
Eine Forschergruppe um Michael Pittman (Pei et al. 2020) wertete die fossilen Daten von 43 Arten von frühen Vögeln und nächstverwandten Formen aus, die zu den Paraves* zusammengefasst werden, um ein aktualisiertes Cladogramm* zu erstellen. Außerdem untersuchten sie anhand einer biomechanischen Analyse das Potenzial dieser Arten zum aktiven Flug. Dazu wurden Körpermerkmale wie Flügelgröße, Körpergröße oder mutmaßliches Lauftempo ausgewertet, um Flächenbelastung der Flügel und den Auftriebskoeffizienten* abzuschätzen.2 Es stellte sich heraus, dass aktive Flieger so unsystematisch im Cladogramm verteilt sind, dass eine mindestens dreimalige Entstehung des aktiven Fluges angenommen werden muss – bei den Vögeln (einschließlich dem berühmten „Urvogel“ Archaeopteryx und der noch etwas älteren Gattung Anchiornis), beim Dromaeosauriden Microraptor (Abb. 1), der relativ klein war und vier Flügel besaß, und bei Rahonavis ostromi (Abb. 2), der lange Vorderextremitäten besaß und einer anderen Linie der Dromaeosauriden angehört (Abb. 3; zu Rahonavis siehe auch Forster et al. [2020]).
Die anderen, nicht zum aktiven Flug befähigten, untersuchten Arten besaßen Merkmale, die ihnen vermutlich das Flattern beim Springen und Laufen, das Gleiten und andere mutmaßliche Vorstufen des aktiven Fliegens ermöglichten.
Evolutionstheoretische Deutungen
Pei et al. (2020) interpretieren die Befunde wie folgt: Viele Vorfahren der nächsten Vogelverwandten hätten sich den Schwellenwerten des Potenzials des aktiven Fluges genähert.3 Das deute auf ein breites „Experimentieren“ mit flügelunterstützter Fortbewegung hin, bevor sich der Flug unter den Dromaeosauriden und Vögeln entwickelt habe. Dazu gehörten schnelles Laufen und Starten, Springen, schnelles Bremsen und Wenden und Steuerung des Gleichgewichts (Pei et al. 2020, 11). Die nicht flugfähigen Formen unter den Paraves sollen gleichermaßen in den Startlöchern gewesen sein.
Dieser Befund unterstütze des Weiteren die Hypothese, dass Schwungfedern und Flügel zuerst für andere Zwecke als für den Flug entwickelt wurden, z. B. andere Arten flügelunterstützter Fortbewegung, Signalgebung („display“) oder das Brüten (Pei et al. 2020, 7).
Glossar
Auftriebskoeffizient: dimensionsloser Koeffizient für den dynamischen Auftrieb eines von einem Fluid umströmten Körpers. Cladogramm: Baumdiagramm, Ähnlichkeitsbaum mit gabeliger Verzweigung; auch als Dendrogramm bezeichnet. Konvergenz: gleichartiges Auftreten ähnlicher Strukturen und Funktionen einzelner Organe nicht abstammungsmäßig verwandter Organismen; evolutionstheoretisch als unabhängige Entwicklung hin zu gleichartigen Merkmalen interpretiert. Paraves: Vögel und ihre evolutionstheoretisch nächstverwandten Gruppen: Üblicherweise werden die Deinonychosaurier (Troodontidae und Dromaeosauridae) dazu gerechnet.
Kritische Anmerkungen und alternative Deutungen
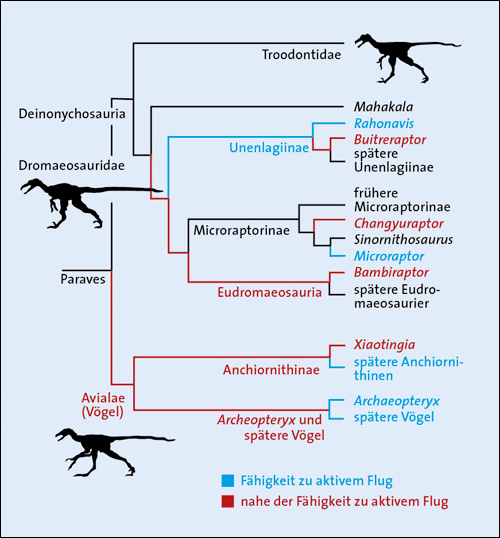
Abb. 3: Teil des Cladogramms von Pei et al. (2020). Die als aktive Flieger rekonstruierten Gattungen stehen auf verschiedenen Ästen des Cladogramms.
Es hat sich eingebürgert, bei Szenarien über größere evolutionäre Übergänge von „Experimentieren“ zu sprechen. Dieser Begriff wird auch von Pei et al. (2020) unkritisch verwendet. Experimentieren ist aber eine bewusste Tätigkeit, durch die man Sachverhalte prüft oder Prozesse optimiert. Ein System dagegen experimentiert nicht; die Befunde scheinen aber ein dem „Experimentieren“ entsprechendes „Prinzip“ nahezulegen. Daher ist dieser Begriff evolutionstheoretisch höchst problematisch, weil er eine Zielorientierung und einen Experimentator impliziert, was für evolutionstheoretische Erklärungen tabu ist. Man muss sich vor Augen halten: Selektion kann erst dann greifen, wenn eine neue Funktion – hier aktiver Flug – wenigstens in schwacher Ausprägung bereits vorhanden ist. Solange das nicht der Fall ist, wird auf andere, (hypothetisch) bereits vorhandene Eigenschaften selektiert, seien es andere Arten von Fortbewegung, Signalgebung oder Brüten, jedoch nicht auf die Fähigkeit zum aktiven Flug. Außerdem ist völlig unklar, warum bei einer Selektion auf Signalgebung oder auf Unterstützung beim Brüten überhaupt flächige Federn mit Detailstrukturen entstehen sollten, die zufällig auch für den aktiven Flug nützlich sind. Aber auch die Entstehung flächiger Federn zur Unterstützung von schnellem Laufen, von Springen oder flügelunterstütztem Klettern würde einen enormen evolutiven Schritt erfordern, der ausgesprochen unplausibel ist, und für die speziellen Eigenschaften der Federn und ihrer Verankerung, die für aktiven Flug erforderlich sind, gäbe es keinerlei Selektionsdruck (detaillierte Diskussion dazu bei Junker [2017]).
Eine Zielorientierung und ein Experimentator sind für evolutionstheoretische Erklärungen tabu.
Welche anderen Deutungsmöglichkeiten der Befunde sind möglich? Einige Wissenschaftler diskutieren im Rahmen der Evolutionstheorie die Möglichkeit, dass die flugunfähigen, aber befiederten Dromaeosauriden sekundär flugunfähig sein könnten; so neuerdings erneut Feduccia (2020). Dafür gibt es einige anatomische Hinweise, aber diese Deutung widerspricht dem Prinzip der Sparsamkeit beim Erstellen von Cladogrammen, d. h. Cladogramme mit der geringsten Anzahl von Konvergenzen werden bevorzugt. Zwar gäbe es in Bezug auf die Flugfähigkeit weniger Konvergenzen; dies würde aber durch eine größere Anzahl an Konvergenzen bei anderen Merkmalen erkauft. Daher wird die Deutung eines sekundären Flugverlusts von der großen Mehrheit der Wissenschaftler abgelehnt. Allerdings sind Cladogramme nicht mit den tatsächlichen Abstammungsverhältnissen gleichzusetzen, sondern es handelt sich um eine Deutung, dass sie bestimmte Abstammungsverhältnisse widerspiegeln. Cladogramme geben nur Ähnlichkeitsverhältnisse wieder, die nach bestimmten Regeln ermittelt wurden und die unabhängig von Ursprungsmodellen ermittelt werden können. Feduccia weist darauf hin, dass die Cladogramme der Paraves ohnehin mit Konvergenzen übersät sind und dass Evolution nicht den sparsamsten Wegen folgen müsse.
Folgt man Feduccia, werden die evolutionstheoretischen Probleme der Entstehung des Vogelfluges aber nicht kleiner, denn der Schritt von nichtfliegenden Vorfahren zu den Vögeln ist noch größer, wenn man nicht annimmt, dass Federn zunächst einen anderen Zweck erfüllt haben. Im Rahmen der Evolutionstheorie geraten alle Ansätze in ausweglose Widersprüche; zum einen wegen der stark mosaikartigen Verteilung der Merkmale und daraus folgend der enormen Anzahl von anzunehmenden Konvergenzen (vgl. Junker 2019), zum anderen wegen des Fehlens plausibler Selektionsdrücke und großer Sprünge, die bewältigt werden müssen, um zu einer neuen Funktion (insbesondere das aktive Fliegen) zu gelangen.
Ein Schöpfer ist frei, sehr unterschiedliche Merkmalskombinationen bei verschiedenen Grundtypen zu verwirklichen.
Aus der Sicht der Schöpfungslehre. Diese Widersprüche kann man im Rahmen eines Schöpfungsansatzes vermeiden. Die Existenz ausgeprägter Mosaikformen und zahlreicher unsystematisch verteilter gemeinsamer Merkmale (was sich in Cladogrammen in Konvergenzen zeigt) ist für Schöpfungsansätze unproblematisch. Denn ein Schöpfer ist frei, sehr unterschiedliche Merkmalskombinationen bei verschiedenen Grundtypen zu verwirklichen, solange die jeweiligen Konstellationen lebensfähige Formen ermöglichen; Evolution ist hingegen an einen natürlichen Mechanismus gebunden. Außerdem entfällt die Suche nach Selektionsdrücken, die ohnehin meistens ad-hoc-Annahmen entspringen und spekulativ sind.
Der Befund der unsystematischen Verteilung von zu aktivem Flug befähigten Formen ist daher im Rahmen eines Schöpfungsansatzes problemlos deutbar. Offenbar gab es zu der Zeit, als die geologischen Schichten von Oberjura und Unterkreide gebildet wurden, in denen die hier in Rede stehenden Organismen eingebettet und fossilisiert wurden, eine große Vielfalt verschiedener Mosaikformen, deren Ähnlichkeitsbeziehungen sich eher netzförmig als baumförmig darstellen.
1 „[i]f independent evolution of key characters is common, how is phylogeny to be reconstructed?“
2 Bei Fossilien muss dabei auf Modellierungen zurückgegriffen werden; direkt bestimmbar sind diese Größen natürlich nicht.
3 „Zum ersten Mal stellen wir fest, dass ein breites Spektrum dieser Vorfahren sich den Schwellenwerten für die Flächenbelastung der Flügel und den Auftriebsbeiwert nähert, die auf ein aktives Flugpotenzial hinweisen“ (Pei et al. 2020, 1).
Romancing the birds and dinosaurs. Irvine, Boca Raton.
The osteology of the Late Cretaceous paravian Rahonavis ostromi from Madagascar. Palaeontologia Electronica, 23(2):a31. https://doi.org/10.26879/793.
Dino-Federvieh – Zum Ursprung von Vogelfeder und Vogelflug. https://www.wort-und-wissen.org/wp-content/uploads/b-17-1_feder-und-flug.pdf.
Sind Vögel Dinosaurier? Eine kritische Analyse fossiler Befunde. https://www.wort-und-wissen.org/wp-content/uploads/b-19-4_dinos-voegel.pdf.
Potential for powered flight neared by most close avialan relatives, but few crossed its thresholds. Curr. Biol. 30, 1–14.
Independent Origins of Middle Ear Bones in Monotremes and Therians. Science 307, 910–914.
Evolutionary cut and paste. Nature 394, 12–13.
Themen | Kurzbeiträge | Streiflichter
Studiengemeinschaft WORT und WISSEN e.V.
Letzte Änderung: 11/24/21
Webmaster