Themen • Kurzbeiträge • Streiflichter
Fossilien widersprechen dem Biogenetischen Grundgesetz
von Henrik Ullrich
Studium Integrale Journal
28. Jahrgang / Heft 2 - September 2021
Seite 110 - 113
Zusammenfassung: In wiederentdeckten Fischfossilien der Gattung Aetheretmon und anderen Gattungen, die als stammesgeschichtliche Vorfahren moderner Fische gelten, fanden sich auch gut erhaltene larvale Stadien. Überraschenderweise unterscheiden sich die ontogenetischen* Entwicklungsschritte der auf 350 Millionen radiometrische Jahre datierten Formen im Bereich der Schwanzentwicklung nicht von denen bei modernen Knochenfischen. Der ontogenetische Entwicklungsweg der Schwanzentwicklung moderner Fische gilt damit nicht mehr als ein Beleg für die Richtigkeit der Rekapitulationsidee von Ernst Haeckel, wonach die Individualentwicklung die Stammesgeschichte (Evolution) wiederholt.
• • • • • •
Ernst Haeckels Biogenetisches Grundgesetz
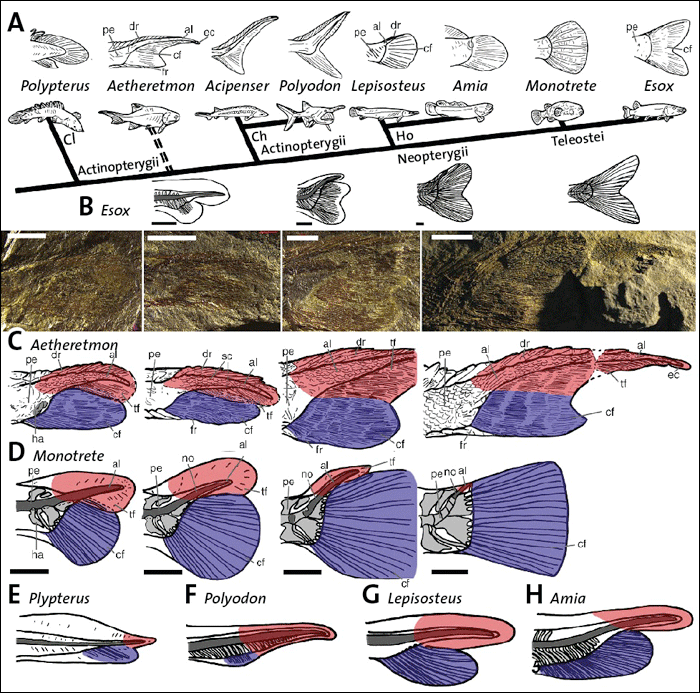
Abb. 1: A Schematische Darstellung der hypothetischen Abfolge evolutionärer Entwicklungsschritte der Schwanzentwicklung der Knochenfische.
B Korrelierend zu A schematische Darstellung der Entwicklung von Schwanzanlagen moderner Fischlarven (Esox, Hecht).
C Fossile Reihe und schematische Darstellung der Schwanzanlagen der fossilen Fischlarven von Aetheretmon.
D Schematische Darstellung der Schwanzanlagen der Fischlarven des heutigen Kugelfisches (Monotrete).
E Schematische Darstellung der Schwanzanlagen von Fischlarven ausgestorbener Formen, die als Vorfahren moderner Fische gelten.
Weitere Erläuterungen siehe Text. (Mit freundlicher Genehmigung aus Sallan 2016)
1866 formulierte Ernst Haeckel (1834–1919) sein Biogenetisches Grundgesetz, das er als naturgesetzliche Grundlage für die Richtigkeit der Abstammungslehre Darwins proklamierte (Haeckel 1866). Dieses Gesetz besagt: 1. Die frühen Stadien von Wirbeltierembryonen weisen eine große Ähnlichkeit mit ihren mutmaßlichen stammesgeschichtlichen Vorfahren auf. 2. Die Kette der aufeinander folgenden embryonalen und fetalen (= ontogenetischen) Bildungsstadien entsprechen im Wesentlichen dem eigenen Formenwandel des Organismus während der Stammesgeschichte, also ihrer Evolution (Haeckel 1866). Das heißt, der Ablauf der Ontogenese eines Individuums ist im Wesentlichen eine kurze und schnelle Wiederholung seiner eigenen evolutionären Geschichte.
Mit einem Stern* versehene Begriffe werden im Glossar erklärt.
Sehr schnell erfuhr dieses „Gesetz“ massive wissenschaftliche Kritik. Bereits Anfang des 20. Jahrhunderts galt die Lehre, die auch unter dem Begriff der Rekapitulation bekannt wurde, im Wesentlichen als widerlegt (siehe u. a. His 1875, Gould 1977; Gursch 1980; Ullrich 1997; 1998; Richardson 1997). In abgewandelter Form, so z. B. unter der Bezeichnung „Biogenetische Grundregel“, wurden die Grundgedanken Haeckels jedoch in verschiedenen Sparten der evolutionsbiologischen Forschung bei der Klärung evolutionärer Entwicklungsschritte von Organen immer wieder genutzt – insbesondere dann, wenn Fossilien fehlten, um die Entstehungsweise eines Organs oder evolutionäre Zwischenstufen hypothetisch zu rekonstruieren (zum Beispiel bei der Evolution des Auges, Kutschera 2006).
Ein in der Fachwelt häufig zitiertes Beispiel für die Gültigkeit von Haeckels Grundideen betrifft die evolutionäre Ausbildung des Schwanzes der echten Knochenfische (Teleostei, z. B. Metscher & Ahlberg 2001). Bisher bekannte adulte* fossile Zwischenformen, welche Hinweise auf die evolutionäre Entstehung der Schwanzstrukturen bei Fischen und bei den Landwirbeltieren gaben, passten gut zu embryonalen Befunden ganz im Sinne von Haeckels Rekapitulationsidee (vgl. Abb. 1A und B). Nun sind es aber gerade wiederentdeckte Fossilien von ausgewachsenen Fischen und ihren Fischlarven aus verschiedenen Entwicklungsstadien aus dem frühen Karbon in Schottland, deren radiometrisches Alter mit 350 Millionen Jahren angegeben wird, welche die bisherigen Thesen widerlegen (Sallan 2016). Die Fossilien werden u. a. der Gattung Aetheretmon zugeordnet (Abb. 1C), einer Gruppe sogenannter ursprünglicher Strahlenflosser, die als evolutionäre Stammform der modernen echten Knochenfische angesehen wird.
Knochenfische: Systematik und evolutionäre Hypothesen
Echte Knochenfische (Teleostei) sind eine Teilklasse der Strahlenflosser (Actinopterygii), die ihrerseits eine Klasse der Knochenfische (Osteichthyes) repräsentieren. Bekannte Vertreter der echten Knochenfische sind Süßwasserfische wie Karpfen oder Barsch oder Meeresfische wie Dorsch oder Hering. Die echten Knochenfische weisen eine riesige Formenfülle auf und sind weltweit in allen Lebensräumen, die Meere, Flüsse und Seen anbieten, zu Hause.
Die vermutete evolutionäre Entwicklung der echten Knochenfische beginnt bei paläozoischen Strahlenflossern vor 350 Millionen Jahren. Diese Fische, wie Polypterus und Aetheretmon, wiesen im ausgewachsenen Stadium einen zweiteiligen schuppenbedeckten Schwanz auf, bestehend aus einem oberen Anteil, der über der eigentlichen unteren Schwanzflosse angeordnet war (vgl. Abb. 1A). Der obere Anteil, welcher aus der Verlängerung der Wirbelsäule hervorgeht, gilt als evolutionäre Anlage des Schwanzes bei Wirbeltieren. Im Verlauf der Stammesgeschichte zu den echten Knochenfischen (Teleostei) habe der untere Anteil den oberen Anteil vollkommen zurückgedrängt bzw. überwachsen und deren typische Schwanzflosse hervorgebracht (z. B. beim Kugelfisch – Monotrete, Hecht – Esox, vgl. Abb. 1A). Die einzelnen Schritte scheinen durch Fossilien belegt zu werden, die in der Reihe über Polyodon, Lepisosteus, Amia zu Monotrete angeordnet das Überwachsen der unteren Anlage dokumentieren.
Dagegen sei bei den Landwirbeltieren, die evolutionär aus der Gruppe der Fleischflosser, den Coelacanthiformes (Quastenflossern) abgeleitet werden, genau das Gegenteilige geschehen. Im Verlauf der Stammesgeschichte habe der obere Schwanzanteil den unteren Anteil zurückgedrängt und durch eine zunehmende Verlängerung den typischen Wirbeltierschwanz hervorgebracht.
Der aktuelle ontogenetische Befund
Larvale Stadien moderner Knochenfische zeigen anatomisch eine Zweiteilung der Schwanzanlage in einen oberen Anteil in der Verlängerung der Wirbelsäule und einen unteren Anteil (vgl. Abb. 1B). Der untere Anteil zeigt während der Ontogenese eine gegenüber der oberen Anlage überproportionale Größenzunahme und bildet schließlich die Schwanzflosse der ausgewachsenen Fische vollständig aus (vgl. auch Abb. 1D beim Kugelfisch – Monotrete). Die Ontogenese der Schwanzstrukturen der modernen Knochenfische ließ sich bisher gut mit den bis dahin vorhandenen fossilen Belegen korrelieren. Die Ontogenese diente ganz im Sinne Haeckels als plausibler Beleg für die vorgeschlagene evolutionäre Transformationsreihe der Schwanzentwicklung.
Glossar
adult: erwachsen, ausgereift, voll entwickelt. Heterochronie: beim Vergleich von Entwicklungssequenzen verschiedener Tierarten nachweisbare Unterschiede hinsichtlich der Reihenfolge des Erscheinens bzw. der Dauer des Nachweises von embryonalen oder ontogenetischen Merkmalen (z. B. der Herzanlage bei Säugetieren und Reptilien). Ontogenese: Individualentwicklung von Organismen vom Zeitpunkt der Befruchtung bis zur Geburt bzw. dem Schlüpfen aus dem Ei; unterteilt in Blastogenese, Embryogenese, Fetogenese. paläozoisch: Das sogenannte „Erdaltertum“ betreffend, dieses umfasst die geologischen Formationen von Kambrium bis Perm. Phylogenese: Theorie der stammesgeschichtlichen Herkunft aller heutigen Lebensformen durch kontinuierliche Abwandlung von Vorgängerarten auf der Grundlage natürlicher evolutiver Prozesse.
Fossile Fischlarven widerlegen Haeckels Biogenetisches Grundgesetz
Lauren Sallan entdeckte nun aber in Sammlungen des Naturhistorischen Museums in London Fossilien ausgewachsener Individuen von Strahlenflossern und bis dahin nicht berücksichtigter Larven dieser Fische. Insbesondere die Funde von Aetheretmon ließen eine nahezu lückenlose Rekonstruktion der larvalen Entwicklung dieses Fisches zu (vgl. Abb. 1B und C). Bei der Analyse kommt die Forscherin zu einem unerwarteten und überraschenden Ergebnis: „Bei unserem Fossil sehen die Entwicklungsschritte aber genauso aus wie bei modernen Fischen. Es kann aber keine Wiederholung geben, wenn es noch nichts zu wiederholen gibt. Man sieht dieselben Entwicklungsschritte bei Fischen vor 350 Millionen Jahren und bei Fischen, die heute heranwachsen“ (zitiert nach Stang 2016).
Auch andere fossile Funde von Fischlarven (z. B. von Polypterus, Polyodon, Lepisosteus oder Amia, vgl. Abbildung E) zeigen bereits die moderne doppelte ontogenetische Anlage des Schwanzes.
„Bei unserem Fossil sehen die Entwicklungsschritte genauso aus wie bei modernen Fischen.“
Für die Autorin legen diese Befunde nahe, dass Veränderungen ontogenetischer Wachstumsprogramme (wie Heterochronie*) die Diversität und Vielfalt des morphologischen Wandels während der Phylogenese bestimmen: „Zusammenfassend lässt sich sagen, dass fossile und ontogenetische Befunde darauf hindeuten, dass die Vielfalt der Wirbeltierschwänze aus einer Heterochronie im Wachstum von zwei verschiedenen Strukturen resultiert, die bei Tetrapoden und Teleostiern aus ähnlichen funktionellen Gründen in entgegengesetzte Richtungen gehen“ (Sallan 2016).
Wieder einmal bestätigt sich, dass die Ontogenese – auch bei Betrachtung von einzelnen Organen – keine Wiederholung der Stammesgeschichte sein kann. „Die Ontogenese rekapituliert nicht die Phylogenese*; stattdessen bestimmen deren differenzierte Entwicklungsschritte die endgültige Morphologie“ (Sallan 2016).
Diskussion
Die Arbeit von Sallan liefert ein weiteres Beispiel zur Widerlegung der Grundideen des Biogenetischen Grundgesetzes von Haeckel. Auch Embryonen fossiler Organismen zeigen unerwartet einen den modernen Organismen ähnlichen Entwicklungsablauf und sind auch deshalb mitnichten als primitiv zu bezeichnen. Dieser Aspekt bestätigt die Ansicht der Schöpfungsforschung, welche von der Vollkommenheit der Organismen seit dem Beginn ihrer Existenz ausgeht.
Embryonen fossiler Organismen zeigen einen den modernen Organismen ähnlichen Entwicklungsablauf und sind mitnichten als primitiv zu bezeichnen.
Auch wenn dieser Aspekt nicht im Zentrum der Darlegungen der Forscherin stand: Die Frage nach den molekularbiologischen bzw. ontogenetischen Mechanismen und den selektiven Rahmenbedingungen der morphologischen Transformation der Schwanzanlagen bei Fischen und Landwirbeltieren bleibt unbeantwortet. Inwieweit die Zuflucht in alternative Evolutionstheorien weiterhilft, wonach insbesondere ontogenetische Mechanismen als maßgebliche Triebfeder des evolutionären Wandels angesehen werden, ist nach wie vor offen.
In manchen Lehrbüchern finden sich immer noch Schemata von Embryonenbildern, die an die alten Abbildungen von Haeckel erinnern. Dabei ist auffällig, dass die Schwanzanlage der Fische in identischer Form auch bei den Landwirbeltier-Embryonen erscheint, egal ob es sich um ein Huhn, einen Affen, ein Schwein oder einen Menschen handelt. Diese Darstellung Haeckels geht davon aus, dass der bei Knochenfischen angelegte embryonale Schwanz als Ganzes der Vorläufer der Schwänze heutiger landlebender Wirbeltiere sei. Unter Berücksichtigung fossiler und ontogenetischer Befunde ist auch dieses Detail seiner Abbildungen aus evolutionärer Perspektive völlig falsch.
Ontogeny and phylogeny. Cambridge.
Die Auseinandersetzungen um Ernst Haeckels Abbildungen. Diss. med. Marburg 1980. Marburger Schriften zur Medizingeschichte. Bern, Frankfurt a.M.
Generelle Morphologie. I: Allgemeine Anatomie der Organismen. II: Allgemeine Entwickelungsgeschichte der Organismen. Berlin.
Unsere Körperform und das physiologische Problem ihrer Entstehung.
Evolutionsbiologie. Stuttgart.
Origin of the teleost tail: phylogenetic frameworks for developmental studies. In: Ahlberg PE (ed) Major Events in Early Vertebrate Evolution. CRC Press, Boca Raton, pp 333–349.
There is no highly conserved embryonic stage in the vertebrates: implications for current theories of evolution and development. Anat. Embryol. 196, 91–106.
Fish ‘tails’ result from outgrowth and reduction of two separate ancestral tails. Curr. Biol. 26, R1205–R1225.
Fossilen stellen „Biogenetische Grundregel“ in Frage. https://www.deutschlandfunk.de/wo-haeckel-sich-irrte-fossilen-stellen-biogenetische.676.de.html?dram:article_id=373475; abgerufen am 22. 8. 2021.
Zur Geschichte der Entdeckung der sogenannten Kiemenbogen und Kiemenspalten in der menschlichen Embryonalentwicklung. Diss. med. Dresden.
Die Wiederentdeckung eines Irrtums. Stud. Integr. J. 5, 3-6.
Themen | Kurzbeiträge | Streiflichter
Studiengemeinschaft WORT und WISSEN e.V.
Letzte Änderung: 11/24/21
Webmaster