Themen • Kurzbeiträge • Streiflichter
Homo naledi einzigartig – aber ein Mensch?
von Michael Brandt
Studium Integrale Journal
28. Jahrgang / Heft 2 - September 2021
Seite 121 - 125
Zusammenfassung: Homo naledi wurde anhand umfangreichen fossilen Materials erstmals 2015 beschrieben. Der frühe Hominine* besitzt ein einzigartiges Merkmalsmosaik bestehend aus einmaligen, einigen menschenähnlichen und zahlreichen großaffenähnlichen Merkmalen. Neue Untersuchungsergebnisse bestätigen dieses Bild. Trotz des menschenuntypischen Merkmalsmosaiks wird die Zuordnung von naledi zur Gattung Homo unter den Paläanthropologen nicht kritisch diskutiert. Homo naledi sollte aber aufgrund seiner Merkmale aus der Gattung Homo ausgegliedert und zu den nichtmenschlichen Homininen in eine eigene Gattung gestellt werden.
• • • • •
Einleitung
Mit einem Stern* versehene Begriffe werden im Glossar erklärt.
Homo naledi ist ein Hominine, der erstmals 2015 anhand von Überresten aus der Dinaledi-Kammer im Rising Star-Höhlensystem in Südafrika und später aus einer zweiten Kammer dieses Höhlensystems, der Lesedi-Kammer, beschrieben wurde. Dieser Hominine weist eine Kombination von Merkmalen auf, die bei keinem anderen Taxon zu finden ist. Versuche, die (vermutete) phylogenetische* Position von Homo naledi zu bestimmen, haben sich als sehr schwierig erwiesen.
Homo naledi besitzt menschenähnliche Merkmale an der Hand und insbesondere am Fuß, zahlreiche großaffenähnliche Merkmale, z. B. an Schulter, Brustkorb, Becken, Hand- und Fußgliedern sowie einzigartige Merkmale, wobei hier die Morphologie* des Daumenmittelhandknochens hervorzuheben ist. Seit der Übersichtspublikation von Brandt (2017) wurden weitere Arbeiten zu Homo naledi publiziert, die die Einzigartigkeit dieses Homininen bekräftigen.
Auch in den jüngsten Studien wird nicht diskutiert, ob die Datenlage die Zuordnung von Homo naledi zum Menschen rechtfertigt. Zu dieser Frage wird am Ende des Beitrages Stellung bezogen. Im Folgenden wird daher der Gattungsname in Anführungszeichen gesetzt.
Neuere Studienergebnisse zu „Homo“ naledi
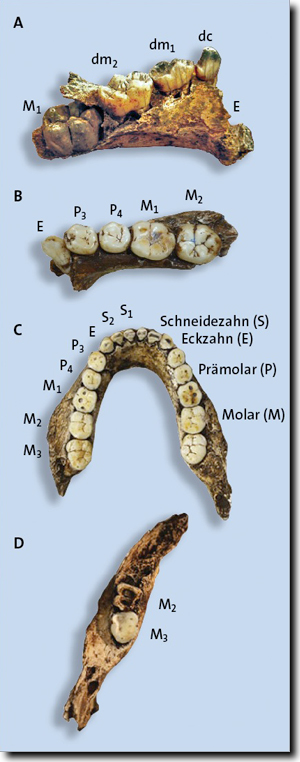
Abb. 1: A Linker Teilunterkiefer eines Kindes mit Milchzähnen und noch nicht durchgebrochener M1-Krone (U.W.101-1400).
B Rechter Teilunterkiefer eines älteren Jugendlichen mit durchgebrochenen bleibenden Zähnen (U.W.101-377).
C Vollständiger Unterkiefer eines erwachsenen Individuums mit allen bleibenden Zähnen (U.W.101-1261).
D Linker Teilunterkiefer eines alten Individuums mit sehr abgenutzten bleibenden Zähnen (U.W.101-361).
Die Abbildungen sind nicht maßstabsgetreu. (Aus Bolter et al. 2018, CC BY 4.0)
Zähne
In Abb. 1 sind vier Unterkiefer mit Zähnen von „Homo“ naledi dargestellt.
Brophy et al. (2021) untersuchten die Kronenform und die Höckerfläche des ersten und zweiten Milchmolaren* von „Homo“ naledi. Dabei waren die Zähne des Unterkiefers diagnostisch deutlich relevanter als die des Oberkiefers.
Die Milchmolaren des Unterkiefers von „Homo“ naledi zeigten in dieser Studie keine nähere Beziehung zu den zum Vergleich herangezogenen zahlreichen menschlichen und nichtmenschlichen Homininen-Taxa (Australopithecus africanus, A. afarensis, Paranthropus robustus, P. boisei, früher Homo sp.1, H. erectus, frühe H. sapiens, jungpaläolithischer H. sapiens, rezenter H. sapiens aus dem südlichen Afrika und Neandertaler). Nach Brophy et al. (2021) stützt das Fehlen einer klaren morphologischen Verwandtschaft und das einmalige Muster der Kronenform und der Höckerfläche der Molaren die Zuordnung von „Homo“ naledi zu einer einzigartigen Art. Frühere Zahnanalysen von „Homo“ naledi haben zu einem ähnlichen Ergebnis geführt (siehe Brandt 2017).
Baily et al. (2019) untersuchten ebenfalls Milchzähne (Schneidezähne, Eckzähne und Molaren) von „Homo“ naledi und verglichen diese auch mit den entsprechenden Zähnen zahlreicher menschlicher und nichtmenschlicher Homininen. Die Milchzähne weisen auch nach dieser Studie eine einmalige Kombination von Merkmalen mit einem Variationsmuster ähnlich dem der bleibenden Zähne auf.
Eine Analyse von Irish et al. (2018) hat wie schon frühere Studien aufgezeigt, dass eine Anzahl von Merkmalen der Kronen und Wurzeln der bleibenden Zähne von „Homo“ naledi auch beim afrikanischen Homo1 zu finden ist. In der Gesamtschau ergibt sich aber ebenfalls ein einmaliges Merkmalsmuster.
Die Zähne von „Homo“ naledi weisen ein Merkmalsmosaik auf, das sich von dem aller anderen Homininen unterscheidet.
Kupczik et al. (2019) untersuchten die Wurzeln der Unterkiefermolaren von sechs Individuen von „Homo“ naledi anhand von fünf metrischen Variablen. In Größe und Gesamtaussehen ähneln die Molarenwurzeln von „Homo“ naledi drei Funden von Homo sp. aus Südafrika. Das Verhältnis des Halses zu den Zweigen der Zahnwurzeln von „Homo“ naledi ist ähnlich dem von Australopithecus africanus und Paranthropus-Arten. „Homo“ naledi teilt auch die Größenfolge des Wurzelvolumens mit Australopithecus und Paranthropus (M2 > M3 > M1), nicht aber mit irgendeiner anderen Homo-Art (M2 > M1 > M3).
Zusammenfassend ist festzustellen, dass alle jüngeren Studienergebnisse der Zähne von „Homo“ naledi Merkmale festgestellt haben, die diese Homininen-Art von allen anderen Homininen deutlich unterscheidet.
Davis et al. (2020) untersuchten die Kronenmorphologie der Prämolaren des Unterkiefers von „Homo“ naledi im Bereich der Schmelz-Dentin-Grenze. Bisherige Studien haben gezeigt, dass sie spezifisch für Homininen-Taxa ist. Die Schmelz-Dentin-Grenze der Prämolaren von „Homo“ naledi unterscheidet sich von der aller anderen Homininen-Taxa. Aus diesem Ergebnis schließen Davis et al. (2020) auf eine Nahrungsnische von „Homo“ naledi in Südafrika, die sich von der aller anderen Homininen unterscheidet.
Auch Berthaume et al. (2018) schließen auf die Bewohnung einer speziellen ökologischen Nische durch „Homo“ naledi. Die Zähne von „Homo“ naledi zeigen, dass dieser frühe Hominine Nahrung mit zwar ähnlichen mechanischen Brucheigenschaften, aber mit mehr abrasiven Partikeln (d. h. abschleifend, z. B. Körner) als Australopithecus, Paranthropus und Homo sp. konsumierte.
Als Retzius-Streifen oder Perikymatien werden die in starker Vergrößerung sichtbaren, über den gesamten Schmelzmantel verteilten Wachstumslinien des Zahnschmelzes bezeichnet. Die Perikymatien unterscheiden sich in der Verteilung bei den frühen Homininen-Arten, wobei es aber auch Musterüberlappungen gibt.
Guatelli-Steinberg et al. (2018) untersuchten bleibende Vorderzähne (Schneidezähne, Eckzähne) von „Homo“ naledi und anderen menschlichen und nichtmenschlichen Homininen. Die Studie zeigt, dass das Merkmalsmuster der Retzius-Streifen von „Homo“ naledi von dem der anderen Homininen unterscheidet. Insgesamt weist dieser Befund auf Unterschiede im Zahnwachstum von „Homo“ naledi im Vergleich zu den anderen Homininen hin.
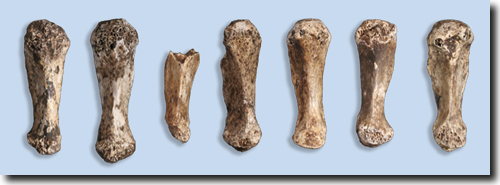
Abb. 2: Daumenmittelhandknochen von „Homo“ naledi in der Ansicht von palmar*. Der Knochen ist einmalig strukturiert. Er weist (oben) eine robuste körperferne Hälfte mit einem mediolateral* weiten Schaft und Kopf und (unten) eine mediolateral und dorsopalmar* schmale proximale Hälfte und Gelenkfläche auf. Auf der Handflächenseite verläuft ein prominenter Kamm. (Aus Berger et al. 2015, CC BY 4.0).
Mittelohr
Elliot et al. (2018) untersuchten den Amboss (Incus) von „Homo“ naledi, der in der Gehörknöchelkette des Mittelohres die mittlere Position zwischen Hammer und Steigbügel einnimmt. Dabei standen den Autoren drei Exemplare zur Verfügung, die wahrscheinlich von drei „Homo“ naledi-Individuen stammen. Die drei Gehörknöchelchen ähneln denen von Schimpansen und Paranthropus robustus mehr als denen vom Menschen (pleistozäner und rezenter Homo sapiens, Neandertaler, Homo aus dem Mittelpleistozän Europas). In der Hauptkomponentenanalyse für größenstandardisierte Variablen liegt „Homo“ naledi innerhalb der Vertrauensellipse für Gorillas. Der Amboss ist somit ein weiterer Knochen unter vielen schon bekannten Skelettelementen von „Homo“ naledi mit einer menschenuntypischen Morphologie.
Hand
Der Daumenmittelhandknochen von „Homo“ naledi (Abb. 2) weist eine sehr eigentümliche Morphologie auf. Dazu gehören ein scharfer medianer Längskamm und ausgeprägte Muskelansätze am Knochen.
Der Daumenmittelhandknochen von
„Homo“ naledi unterscheidet sich von dem der lebenden und fossilen Primaten.
Bowland et al. (2018, 2021) unterzogen diesen Knochen einer erneuten Analyse. Die Autoren kommen zu dem Ergebnis, dass sich „Homo“ naledi von anderen Homininen durch die Kombination aus einer kleinen proximalen* Basis, einem grazilen Schaft und distal* breiten Muskelansätzen (Enthesis) unterscheidet. In diesen Merkmalen ist ein Australopithecus-Fund (StW 418) „Homo“ naledi jedoch ähnlich (Bowland et al. (2021). Der mediane Längskamm des Daumenmittelhandknochens von „Homo“ naledi wurde von den Autoren nicht vergleichend untersucht. Die Grazilität des proximalen Schaftes und der Basis von „Homo“ naledi ist ähnlich wie bei Hundsaffen, Orang-Utans, Schimpansen, Australopithecus afarensis und Australopithecus sediba (Bowland et al. 2018, 2021).
Im Gegensatz dazu zeichnet sich der Daumenmittelhandknochen des Neandertalers und SK 84 (Paranthropus oder Homo erectus) durch eine robuste Basis, einen zusammengedrückten Mittelschaft und einen breiten Kamm (Enthesis) für den Ansatz des Musculus opponens pollicis („Gegensteller-Muskel des Daumens“) aus. Dagegen besitzt der moderne menschliche Daumenmittelhandknochen eine robuste Basis in Kombination mit einem geraden Schaft und einer mäßigen Enthesisausprägung.
Eine Struktur des Daumenmittelhandknochens von „Homo“ naledi ist jedoch menschenähnlich: die distale Gelenkfläche (Galetta et al. 2019).

Abb. 3: Rechtes Schlüsselbein U.W. 102a-021 aus der Lesedi-Kammer. Links Ansicht von (v. o. n. u.) oben, vorn, hinten und unten. Rechts Ansicht von (oben) innen und (unten) außen. (Aus Hawks et al. 2018, CC BY 4.0)
Schulter
Obere Extremitätenknochen von „Homo“ naledi aus der Dinaledi-Kammer wurden bereits
publiziert. Feuerriegel et al. (2018) stellten von dieser Spezies neue obere Extremitätenfossilien aus der Lesedi-Kammer vor. Sie stammen von einem Teilskelett (LES 1). Nach Feuerriegel et al. (2018) zeigt das neue Knochenmaterial aus der Lesedi-Kammer funktionell bedeutsame morphologische Aspekte, die beim Fossilmaterial aus der Lesedi-Kammer fehlen. Zu dem neuen Material gehören ein vollständiges Schlüsselbein, ein gut erhaltener oberer Teil eines Oberarmknochens und der obere Teil einer Elle (Abb. 3-5).
Insgesamt zeigen diese Fossilien wie schon das Material aus der Dinaledi-Kammer Ähnlichkeiten mit Australopithecus. „Homo“ naledi hielt zu Lebzeiten seine Schulter vergleichbar mit höher als der moderne Mensch und war somit an das Baumleben angepasst.
Oberschenkel
Friedl et al. (2018) untersuchten die Struktur des Halses und des Schaftes des Oberschenkelknochens von „Homo“ naledi. Insgesamt zeigt der proximale Femur ein einzigartiges Merkmalsmosaik. Es weist auf ein einmaliges Belastungsmuster hin, das wahrscheinlich in Beziehung zu der einmaligen Anatomie des postcranialen* Skeletts dieser Spezies steht. Nach Friedl et al. (2018) weist das einmalige Merkmalsmosaik des Femurs unzweifelhaft auf eine gewohnheitsmäßig zweibeinige Fortbewegung von „Homo“ naledi auf dem Erdboden hin. Dies ist eine kühne, nicht naheliegende Schlussfolgerung, denn nur der Mensch bewegt sich unter den Primaten gewohnheitsmäßig zweibeinig fort und besitzt in diesem Zusammenhang zahlreiche Merkmale, die „Homo“ naledi nicht aufweist (siehe Brandt 2017).
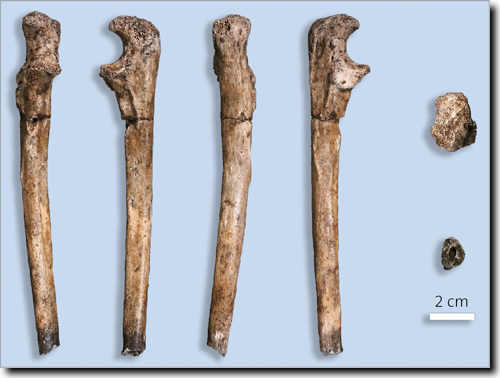
Abb. 4: Rechtes körpernahes Ulna-Fragment U.W. 102a-015 aus der Lesedi-Kammer. Links Ansicht von (v. l. n. r) vorn, innen, hinten und außen. Rechts Ansicht von (oben) körperfern und (unten) körpernah. (Aus Hawks et al. 2018, CC BY 4.0)
Fuß
• Sprungbein. Kasl et al. (2019) untersuchten die Trabekelstruktur* von vier Sprungbeinen von „Homo“ naledi. Die Struktur der Trabekel dieses Knochens, welche mit dem Belastungsmuster bei der Fortbewegung im Zusammenhang steht, ist bei modernen Großaffen, modernen Menschen und fossilen Homininen unterschiedlich. Der Grad der Anisotropie (Vorzugsrichtung) der Trabekel des Sprungbeines von „Homo“ naledi folgt einem Muster ähnlich dem des modernen Menschen. Hierbei ist aber anzumerken, dass die Methode zur Bestimmung der Anisotropie generell ein großes Problem ist.2

Abb. 5: Linkes körpernahes Oberarmknochen-Fragment U.W. 102a-257 aus der Lesedi-Kammer. Links Ansicht von (v. l. n. r.) hinten, innen, vorn und außen. Rechts Ansicht von (oben) körpernah und (unten) körperfern. (Aus Hawks et al. 2018, CC BY 4.0)
Die Trabekeldicke und das Verhältnis von Knochenvolumen zu Trabekelvolumen ist dagegen ähnlich wie bei den lebenden Großaffen und einigen fossilen Homininen. Kasl et al. (2019) schließen aus ihren Befunden auf eine vorherrschend menschenähnliche Belastung des oberen Sprunggelenkes.
• Längenverhältnis Mittelfußknochen/Zehen. Traynor et al. (2018) bestimmten bei „Homo“ naledi das Längenverhältnis der proximalen Phalangen zu den Mittelfußknochen. Die Längenproportion des ersten und zweiten Fußstrahls ist menschenähnlich, die des dritten und vierten Fußstrahls jedoch menschenunähnlich. Die Autoren schließen von diesem Ergebnis auf die Möglichkeit einer einzigartigen Funktion der inneren gegenüber der äußeren Fußsäule.
Der Fuß von „Homo“ naledi besitzt neben menschenähnlichen Merkmalen auch eine ganze Anzahl nichtmenschlicher Anpassungen, die auf eine einmalige Fußfunktion hinweisen.
• Fünfter Mittelfußknochen. Der fünfte Mittelfußknochen von „Homo“ naledi ist in seiner äußeren Form menschenähnlich (DeSilva et al. 2018) und besitzt eine menschenähnlich geringe Dicke der Rinde (Dowdeswell et al. 2017). Allerdings ist die Steifheit dieses Knochens großaffenähnlich und es fehlt eine menschenähnliche Verjüngung nach distal. Mit dieser Merkmalskombination unterscheidet sich „Homo“ naledi von allen anderen Homininen. Dowdeswell et al. (2017) ziehen von diesen anatomischen Verhältnissen den weitreichenden funktionellen Schluss auf eine fehlende menschenähnliche Belastung des Fußes von „Homo“ naledi im Außenbereich (laterale Säule), fügen aber einschränkend hinzu, dass die funktionelle Interpretation der einmaligen Morphologie des fünften Mittelfußknochen von „Homo“ naledi weitere vergleichende Untersuchungen des Körperstamm- und Extremitätenskeletts erfordert.
Wer war „Homo“ naledi?
Die neueren Studien haben das bisherige morphologisch-funktionelle Bild von „Homo“ naledi verfestigt. Dieser fossile Hominine besitzt ein ausgeprägt einzigartiges Merkmalsmosaik aus einmaligen Merkmalen, vielen großaffenähnlichen Merkmalen und einigen menschenähnlichen Merkmalen. Wegen dieses einmaligen Merkmalsmosaiks ist die (angenommene) evolutionäre Stellung von „Homo“ naledi unter Paläanthropologen unklar. So bezeichnen Brophy et al. (2021) die Stellung von „Homo“ naledi als „rätselhaft“ und Baily et al. (2019) als „zweideutig gegenüber anderen Homo-Arten“. Neben dem ausgeprägten einmaligen Merkmalsmosaik trägt auch das unerwartet geringe geologische Alter zu der unklaren Situation in einem evolutionären Rahmen bei. Wegen seiner Merkmale hatte man für „Homo“ naledi ein hohes geologisches Alter vermutet. Stattdessen ist dieser Hominine mit 236.000–335.000 Isotopenjahren nur wenig älter als Homo sapiens mit 200.000 Isotopenjahren. In Anbetracht des einmaligen Merkmalsmosaiks mit zahlreichen großaffenähnlichen Merkmalen ist es unverständlich, warum naledi, der sich von allen unzweifelhaft echten lebenden und fossilen Menschen deutlich unterscheidet, in die Gattung Homo gestellt wurde. Die zahlreichen nichtmenschlichen Merkmale rücken naledi in die Nähe der nichtmenschlichen Homininen. Das von allen Homininen deutlich verschiedene Merkmalsmosaik ist sogar mit einer Einordnung dieser Spezies in eine neue Gattung gut vereinbar. Es ist deshalb verwunderlich, dass die bisherige taxonomische Zuordnung von naledi zu Homo von Paläanthropologen nicht kritisch diskutiert wird.
Im Grundtypkonzept der Schöpfungslehre ist hingegen die Deutung plausibel, dass naledi eine Spezies einer neuen Gattung innerhalb eines Großaffengrundtyps repräsentiert.
Glossar
distal: körperfern. dorsopalmar: von der Rückseite zur Handfläche. Hominine: Menschenähnliche; damit werden alle fossilen und lebenden Menschenformen einschließlich ihrer im Evolutionsmodell vermuteten Vorläufer bezeichnet. mediolateral: von der Mitte zur Seite hin. Milchmolar: Mahlzahn des Milchgebisses. Morphologie: Wissenschaft vom Bau und der Gestalt des Körpers der Lebewesen und seiner Organe. palmar: handflächenseitig. phylogenetisch: im Evolutionsmodell die stammesgeschichtliche Entwicklung aller Lebewesen betreffend. postcranial: dem Skelett mit Ausnahme des Schädels zugehörig. proximal: körpernah. Trabekelstruktur: feine Strukturen aus Knochengewebe (Bälkchen), mit denen der Innenraum vieler Knochen aufgebaut ist.
1 In der Paläanthropologie werden zur Gattung Homo auch Funde gestellt, die keine echten Menschen waren. Deshalb ist bei Fossilien des Taxons Homo sp. unklar, ob es sich um echte Menschen handelt.
2 Shabestari OM (2002) Zur Struktur des knöchernen Implantatlagers nach Morphometrie bioptischer und autoptischer Fälle. FU Berlin: Digitale Dissertation: http://webdoc.sub.gwdg.de/ebook/diss/2003/fu-berlin/2002/271/Kap01.pdf
The deciduous dentition of Homo naledi: A comparative study. J. Hum. Evol. 136, 102655.
Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa. eLife 4:e09560.
Dental topography and the diet of Homo naledi. J. Hum. Evol. 118, 14–26.
Corrigendum. J. Hum. Evol. 131, 240.
Palaeodemographics of individuals in Dinaledi Chamber using dental remains. S. Afr. J. Sci. 114, 1–6.
Pollical metacarpal shaft morphology in Homo naledi: A 3D geometric morphometric analysis. Am. J. Phys. Anthropol. 165, S66, 34.
Homo naledi pollical metacarpal shaft morphology is distinctive and intermediate between that of australopiths and other members of the genus Homo. J. Hum. Evol. 158, 103048.
Homo naledi – neuer Hominine mit vielen Fragezeichen. In: Brandt M: Frühe Homininen. Eine Bestandsaufnahme anhand fossiler und archäologischer Zeugnisse. Studium Integrale Special 1, 51–79.
Comparative morphometric analysis of the deciduous molars of Homo naledi from Dinaledi Chamber, South Africa. Am. J. Phys. Anthropol. 174, 299–314.
Distinct mandibular premolar crown morphology in Homo naledi and its implications for the evolution of Homo species in southern Africa. Scientific Reports 10:13196.
One small step: A review of Plio-Pleistocene hominin foot evolution. Yrb. Phys. Anthropol., 1–78.
Adaptation to bipedal gait and fifth metatarsal structural properties in Australopithecus, Paranthropus, and Homo. C. R. Palevol 16, 585–599.
Description and analysis of three Homo naledi incudes from the Dinaledi Chamber, Rising Star cave (South Africa). J. Hum. Evol. 122, 146–155.
The upper limb of Homo naledi. New material from the Lesedi Chamber, Rising Star System, South Africa. Am. J. Phys. Anthropol. 165, S66, 84.
Femoral neck and shaft structure in Homo naledi from the Dinaledi Chamber (Rising Star System, South Africa). J. Hum. Evol. 133, 61–77.
Three-dimensional geometric morphometric analysis of the first metacarpal distal articular surface in humans, great apes and fossil hominins. J. Hum. Evol. 132, 119–136.
Patterns of lateral enamel growth in Homo naledi as assessed through perkymata distribution and number. J. Hum. Evol. 121, 40–54.
New fossil remains of Homo naledi from the Lesedi Chamber, South Africa. eLife 6:e24232.
Ancient teeth, phenetic affinities, and African hominins: Another look at where Homo naledi fits in. J. Hum. Evol. 122, 108–123.
An analysis of the trabecular morphology of the Homo naledi talus, and its inferred functional implications. Am. J. Phys. Anthropol. 168, S68, 120.
Mandibular molar root und pulp cavity morphology in Homo naledi and other Plio-Pleistocene hominins. J. Hum. Evol. 130, 83–95.
Metatarsophalangeal proportions of Homo naledi. Am. J. Phys. Anthropol. 165, S66, 276.
Themen | Kurzbeiträge | Streiflichter
Studiengemeinschaft WORT und WISSEN e.V.
Letzte Änderung: 11/24/21
Webmaster