
Themen • Kurzbeiträge • Streiflichter
Die „Sauerstoffkatastrophe“
1. Extreme Krise als Auslöser genialer „Neuerfindungen“ des Lebens?
von Boris Schmidtgall
Studium Integrale Journal
28. Jahrgang / Heft 1 - April 2021
Seite 13 - 21
Zusammenfassung: Nach evolutionstheoretischen Vorstellungen sollen die ersten Lebewesen in einer sauerstofffreien Atmosphäre entstanden sein. Denn eine Entstehung der ersten Makromoleküle des Lebens ist bei Vorhandensein von Sauerstoff chemisch nicht möglich. Daher wird angenommen, dass die Umstellung zu einer mit Sauerstoff angereicherten Atmosphäre, wie wir sie heute kennen, erst später im Zuge der postulierten Evolution der Biosphäre erfolgte. Dieses vermutete Szenario einer frühen Evolution wird üblicherweise als Great Oxygenation Event (GOE) bezeichnet. Um diese Hypothese zu stützen, sind viele geologische und paläobiologische Studien durchgeführt worden. Sie ergeben jedoch ein unklares und widersprüchliches Bild von den ersten Schritten des Lebens auf der Grundlage der Darwin’schen Evolutionslehre.
• • • • • •
Mit einem Stern* versehene Begriffe werden im Glossar erklärt.
Einführung
Ein bekanntes Sprichwort lautet: „Not macht erfinderisch.“ Dies gilt durchaus für den Verlauf der Menschheitsgeschichte, denn viele bahnbrechende Erfindungen der Menschheit stammen aus Krisenzeiten. Errungenschaften wie die Landwirtschaft oder die Mühle dienten dazu, den zunehmenden Nahrungsbedarf einer wachsenden Bevölkerung zu decken. Und das Rad, die Metallverhüttung oder das Schießpulver wurden vermutlich erfunden, um Kriege zu gewinnen. Dass Erfindungen durch Beobachtung, gründliche Überlegung, Experimente, Beratung und Planung zustande kommen, sprich durch den Einsatz von Intelligenz, ist so selbstverständlich, dass es eigentlich keiner Erwähnung bedarf. Ungeachtet dessen sprechen Befürworter der Evolutionslehre oft von „Erfindungen“, um die evolutive, d. h. plan- und ziellose Entstehung komplexer biochemischer oder biologischer Funktionseinheiten zu beschreiben. In manchen Fällen wird auch von „Revolutionen“ des Lebens gesprochen, um große, angeblich durch evolutive Vorgänge bewirkte Veränderungen zu beschreiben, wobei auch dieser Begriff in seiner eigentlichen Bedeutung hierfür unpassend ist.
Als die wohl allergrößte „Revolution“ und „Erfindung“ in der Geschichte der Lebewesen würde wahrscheinlich die angenommene evolutive Entstehung sauerstoffproduzierender Organismen gelten. Homann-Marriott & Blankenship (2011) beschreiben diesen hypothetischen Vorgang mit genau diesen Begriffen: „Die Erfindung der sauerstofferzeugenden Photosynthese und der anschließende Anstieg des atmosphärischen Sauerstoffs vor etwa 2,4 Milliarden Jahren haben die energetischen und enzymatischen Grundlagen des Lebens revolutioniert“ (Hervorhebungen hinzugefügt).
Demnach soll es in der Geschichte des Lebens eine Epoche gegeben haben, in der eine vorher nahezu komplett sauerstofffreie Erdatmosphäre mit diesem reaktiven Gas angereichert worden ist. Dieses hypothetische Szenario wird in der Fachliteratur üblicherweise als „Great Oxygenation Event“ (GOE) bezeichnet und gilt als „Rätsel“ oder „Mysterium“, da es sehr viele Fragen aufwirft. Zusammenfassend kommentierten Sessions et al. in ihrem Artikel „Das fortwährende Rätsel der großen Sauerstoffkatastrophe“1 (2009) die Auseinandersetzung mit dieser Frage mit den Worten: „Während der Sauerstoffanstieg Gegenstand einer beträchtlichen Aufmerksamkeit seitens der Geowissenschaftler war, bleiben einige entscheidende Aspekte dieses Problems ungelöst.“
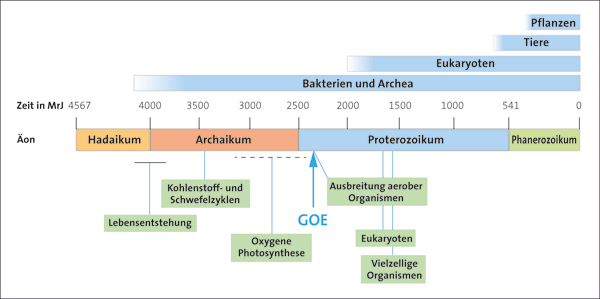
Abb. 1: Zeittafel der hypothetischen Evolution nach Knoll & Nowak (2017). Die Zeit auf der Skala ist in Millionen radiometrischen Jahren (MrJ) angegeben. Als „Äonen“ werden die Zeitabschnitte auf der Skala bezeichnet.
Das GOE gilt in der Fachliteratur als „Rätsel“ oder „Mysterium“, da es sehr viele Fragen aufwirft.
Die Beschreibung dieses Ereignisses wird häufig grob wie in dem folgenden Modell wiedergegeben (Sessions et al. 2009): Die ersten Organismen entstanden vor ca. 4 Milliarden Jahren. Sie lebten in einer sauerstofffreien Atmosphäre und bezogen ihre Energie aus dem Licht oder verschiedenen chemischen Stoffen. Dann entwickelten sich vor ca. 2,4 Milliarden Jahren photosynthetisch aktive Bakterien, die Sauerstoff als Abfallprodukt ihres Stoffwechsels erzeugten. Dies führte dazu, dass die Atmosphäre sich allmählich mit Sauerstoff anreicherte. Dabei sollen zunächst lokale „Sauerstoff“-Oasen entstanden sein und in der Zeit von vor ca. 2,1-1,0 Milliarden Jahren zu einer allmählichen geringfügigen Anreicherung der Atmosphäre mit Sauerstoff geführt haben (ca. 1-2 Vol% der Atmosphäre). Im Verlauf der darauf folgenden 500 Millionen Jahre soll eine durchgreifende, globale Anreicherung der Erdatmosphäre mit Sauerstoff (Oxygenierung) erfolgt sein, so dass etwa der Zustand, in welchem sie heute vorliegt, erreicht wurde (78,08% Stickstoff, 20,95% Sauerstoff, <1% Spurengase: u. a. Kohlendioxid, Methan, Edelgase).2 Die Tiefsee soll jedoch noch bis vor 500 Millionen Jahren überwiegend anoxisch* geblieben und erst später allmählich vom Sauerstoff durchdrungen worden sein. Durch die Konfrontation mit diesem reaktiven Gas sollen die damals lebenden Organismen zu einer immensen Umgestaltung ihrer biochemischen Ausstattung und Lebensweise gezwungen worden sein. Lebensformen, denen das nicht gelungen ist, müssten demzufolge entweder ausgestorben oder in sauerstofffreie Nischen ausgewichen sein. Eine Zusammenfassung des Standardmodells der Evolution in grober zeitlicher Auflösung ist in Abb. 1 dargestellt.
Im Hinblick auf das GOE stellen sich unmittelbar grundlegende Fragen: Welche Indizien gibt es dafür, dass das Leben in einer anoxischen Atmosphäre angefangen haben könnte? Wie hätte Leben in einer Atmosphäre entstehen können, die nicht von einer Ozon-Schicht geschützt wurde und daher lebensfeindlicher harter UV-Strahlung ausgesetzt war? Wie könnten die Stoffkreisläufe (z. B. Kohlenstoffkreislauf, Stickstoffkreislauf etc.) ohne den Sauerstoff funktioniert haben? Gibt es Belege für eine schrittweise Anreicherung der Atmosphäre mit Sauerstoff? Welche Selektionsdrücke und Mechanismen könnten die Entstehung der Photosynthese bewirkt haben, durch die der Sauerstoff erzeugt wird? In mehreren Folgen sollen diese und andere Fragen aufgegriffen werden, um eines der größten „Wunder der Evolution“ auf den Prüfstand zu stellen. In diesem Beitrag geht es um geologische und paläobiologische Indizien, die angeführt werden, um das GOE zu belegen.
In Gegenwart von Sauerstoff können keine Bausteine des Lebens entstehen. Daher ist die Annahme eines anoxischen Lebensanfangs zwingend für die Evolutionslehre.
Ursprung der Idee eines anoxischen Anfangs des Lebens
Die Annahme, dass erste Lebensformen sich in einer sauerstofffreien Atmosphäre gebildet haben sollen, stammt von dem russischen Biochemiker A. I. Oparin (1938/1949). Seine Arbeit beruhte nicht auf experimentellen Ergebnissen, sondern allein auf chemischer Logik. Und diese Logik erfordert aufgrund der Empfindlichkeit der molekularen Bausteine des Lebens gegenüber Sauerstoff, dass die frühe Atmosphäre frei von Sauerstoff und reduzierend* gewesen sein musste, um die Entstehung dieser Moleküle zu ermöglichen. Diese Annahme wurde durch die Ursuppen-Experimente von Miller & Urey (1953) bestätigt. In diesen Experimenten erhielt man Aminosäuren, indem elektrische Entladungen in einer Atmosphäre bestehend aus Methan, Ammoniak, Wasserdampf und Wasserstoff über einen längeren Zeitraum ausgelöst wurden. Zusammenfassend schrieben Miller & Urey später (1959): „Wenn die [experimentellen] Bedingungen oxidierend* waren, entstanden keine Aminosäuren. Diese Experimente haben die Hypothese bestätigt, derzufolge reduzierende Atmosphären für die Entstehung organischer Verbindungen in nennenswerten Mengen erforderlich sind.“ Aufgrund der beschränkten Aussagekraft der Miller‘schen Labor-Experimente bestand großes Interesse, geologische und paläobiologische Hinweise zu erhalten, die Rückschlüsse auf den tatsächlichen Zustand der frühen Erdatmosphäre und das Leben der ersten Organismen ermöglichen.
Kompakt
Gemäß der Darwin’schen Evolutionslehre sollen die ersten Lebewesen durch chemische Vorgänge entstanden sein. Ein solcher Vorgang ist jedoch in Gegenwart von Sauerstoff aufgrund der Empfindlichkeit der molekularen Bausteine des Lebens gegenüber diesem reaktiven Gas nicht möglich. Dieser Sachverhalt war richtungsgebend für die weitere Hypothesenbildung und Forschung bezüglich der Evolution der frühen Organismen. Lebensursprungsforscher gingen daher stets davon aus, dass die ersten Mikroorganismen sich in einer sauerstofffreien Atmosphäre vor ca. 4-3,8 Milliarden Jahren entwickelt haben sollen. Später sollen sauerstoffproduzierende Mikroorganismen evolviert sein, sodass die Atmosphäre sich mit Sauerstoff anreicherte. Dieser Vorgang wird als Great Oxygenation Event (GOE) bezeichnet und soll gemäß konventioneller Vorstellung vor ca. 2,4 Milliarden Jahren eingesetzt haben.
Zur Stützung dieser Annahme werden geologische und paläobiologische Befunde angeführt. Dazu gehören Verteilungen bestimmter Mineralien und Isotopen in den Gesteinsschichten sowie fossile Reste oder molekulare Marker von Mikroorganismen. Insgesamt ergibt die Summe der Befunde in keiner Weise ein klares Bild, das einen tatsächlich stattgefundenen Wechsel von einer sauerstofffreien zu einer sauerstoffreichen Atmosphäre nahelegt. Der Beginn des GOE schwankt je nach Art der Befunde um viele Hundert Millionen Jahre und auch der Verlauf wird entweder als sehr schnell (1-10 Millionen Jahre) oder als sehr langsam eingeschätzt (mehrere Hundert Millionen Jahre).
Darüber hinaus erscheint das gesamte Szenario in vielerlei Hinsicht unplausibel zu sein. Die Abwesenheit von Sauerstoff bedingt, dass die vor harter UV-Strahlung schützende Ozonschicht gänzlich fehlen würde. Zudem würde eine frühe Etablierung eines Kohlenstoffkreislaufs ohne Sauerstoff zwangsläufig zu einer Anhäufung des starken Treibhausgases Methan führen, sodass ein Backofen-Klima auf der Erde geherrscht hätte. Problematisch wäre auch der Umschwung zu einer sauerstoffreichen Atmosphäre, da dann das Methan schnell abgebaut würde und so eine globale Vereisung eingetreten wäre. Wie hypothetische frühe Organismen solche Schwankungen überstanden haben sollen, ist völlig unklar. Nur durch Rückgriff auf spekulative Annahmen kann das Modell aufrechterhalten werden.
Geologische Indizien für das GOE
Als eine erste wichtige Kategorie von Indizien für den angenommenen Umschwung von einer sauerstofffreien hin zu einer mit Sauerstoff angereicherten Atmosphäre werden geologische Indizien angeführt. Anhand von unterschiedlichen Verteilungsmustern bestimmter Mineralien und Isotopen* in den Gesteinen sucht man nach Hinweisen auf eine Änderung der Zusammensetzung der Erdatmosphäre in früheren Epochen der Erdgeschichte.
Vorkommen bestimmter Mineralien

Abb. 2: Amerikanisches Bändereisenerz; die roten „Bänder“ sind angereichert mit Eisen(III)oxid (Fe2O3). (Quelle: farbkombinat, AdobeStock)
Präkambrische Bändereisenerze
Ein gründlich untersuchter geologischer Befund, der als Indiz für das GOE gewertet wird, sind die präkambrischen Bändereisenerze (Banded Iron Formations, BIFs, s. Abb. 2). Dazu werden zwei Schichtfolgen gerechnet, die dem späten Archaikum (>2.5 Milliarden radiometrische Jahre) beziehungsweise dem Paläoproterozoikum (ca. 1,5 – 2,1 Milliarden radiometrische Jahre) zugeordnet werden. Bei den BIFs handelt es sich um flächenmäßig relativ weit ausgedehnte Sedimentfolgen mit Mächtigkeiten von bis zu einigen 100 m (Gole & Klein 1981).
Die Relevanz der Bändereisenerze für den GOE rührt daher, dass sie einen hohen Anteil (ca. 20 bis 40%) an Eisenmineralien enthalten, die zu einem großen Teil als Fe2O3 (Eisen(III)oxid) vorliegen. Das Vorliegen des Eisens in der höchsten Oxidationsstufe (Fe3+) in diesen Schichten wird als möglicher Hinweis auf eine Anreicherung der Atmosphäre mit Sauerstoff (O2) zur Zeit der Ablagerung der BIFs interpretiert. Als Erklärung für die Ablagerung der BIFs werden bevorzugt biologische Ursachen angeführt. Als eine Möglichkeit wurden photosynthetisch aktive Mikroorganismen vorgeschlagen, die Sauerstoff freigesetzt haben sollen, sodass sich lokal „Sauerstoff-Oasen“ bildeten (Cloud 1973; Fralick & Pufahl 2006). Durch die chemische Reaktion von Sauerstoff mit im Wasser gelösten Fe(II)-Ionen soll sich das unlösliche Fe2O3 gebildet und als rötlicher Niederschlag abgesetzt haben.
Andere Forscher schlugen photoferrotrophe* Organismen als Verursacher der Eisen(III)oxid-Ablagerungen vor (Posth et al. 2013). Dabei handelt es sich um Mikroorganismen, die Photosynthese mit Fe(II)-Ionen anstelle von Wasser als Elektronenquelle betreiben. Solche Organismen erzeugen dementsprechend Fe(III)-Ionen als „Abfallprodukt“ anstelle von O2 (Thompson et al. 2019). Bei diesem überaus komplexen biochemischen Gesamtvorgang findet auch CO2-Fixierung unter Aufbau von Biomasse statt. Damit jedoch ein Stoffkreislauf stattfinden kann, muss diese Biomasse später wieder abgebaut und dem Aufbau neuer Mikroorganismen als CO2 zur Verfügung stehen. Da in den BIFs jedoch keine Spuren von abgelagerter Biomasse zu finden sind, schlugen Thompson et al. vor, dass der Abbau der Biomasse durch mikrobielle Methanerzeugung stattfand, sodass die frühe Atmosphäre relativ stark mit Methan angereichert gewesen sein müsste.
Die Annahme von Thompson et al., dass ein Kohlenstoffkreislauf bereits in einer sauerstofffreien Atmosphäre etabliert war, ist durchaus fragwürdig. Denn nach diesem Szenario müssten die methanogenen Mikroorganismen praktisch zur gleichen Zeit wie die photoferrotrophen entstanden sein. Das würde aber einen sehr großen evolutionären Sprung erfordern und nicht zu einer langsam voranschreitenden Evolution nach Darwin’schem Muster passen. Zudem stellt sich die Frage, wie sich die Anreicherung des Methans in der sauerstofffreien Atmosphäre ausgewirkt haben könnte, da Methan als starkes Treibhausgas schon bei relativ kleinen Volumenanteilen in der Atmosphäre ein Backofen-Klima erzeugen kann. Der Treibhauseffekt von Methan beträgt etwa das 25fache desjenigen von CO2.3 Thompson et al. stützen daher ihr Modell mit der Annahme einer im Vergleich zu heute um etwa 20% schwächeren Sonnenintensität, die den Treibhauseffekt kompensiert haben soll: „Folglich würde die Ablagerung von BIFs durch photoferrotrophe [Organismen] dazu beitragen, dass die Atmosphäre sich mit Methan anreichert, sodass das Klima der Erde unter einer trüben frühen Sonne stabilisiert werden könnte.“ Demnach wird eine präzise Feinabstimmung der schwächeren Sonnenintensität und des früheren hypothetischen Kohlenstoffkreislaufs vorausgesetzt – eine rein spekulative Annahme.
Wie mutmaßlich frühe Organismen die vermuteten Schwankungen zwischen Backofen-Klima und globaler Vereisung überstanden haben sollen, ist völlig unklar.
Eine weitere Ad-hoc-Annahme*, auf die sich das Modell der anoxischen Atmosphäre stützt, ist die Kompensation der fehlenden Ozonschicht durch die um 20% geringere Intensität der Sonneneinstrahlung. Denn in Abwesenheit von Sauerstoff konnte es diesen wirksamen Schutz der Lebewesen vor der vernichtenden harten UV-Strahlung nicht gegeben haben. Zudem muss davon ausgegangen werden, dass das spätere Hinzukommen des Sauerstoffs im Zuge des GOE zu einer globalen Vereisung führte, da der Sauerstoff im Gegensatz zu Treibhausgasen einen Wärmeverlust der Atmosphäre und den raschen Abbau des Methans fördert. Manche Autoren nehmen gar von mehreren O2-Schüben ausgelöste, wiederholte globale Vereisungen an, die später durch verstärkte Vulkanaktivität wieder rückgängig gemacht worden sein sollen (Gumsley 2017). Wie das hypothetische frühe mikrobielle Leben, das auf Licht angewiesen gewesen sein soll, diese enormen Schwankungen der Lebensbedingungen hätte meistern können, ist völlig unklar.
Andere Mineralien
Während Eisen(II)-Mineralien durch Reaktion mit Sauerstoff zu kaum wasserlöslichem Fe2O3 umgesetzt werden, gibt es andere Mineralien, die durch Reaktion mit Sauerstoff löslich werden. Das gilt insbesondere für Sulfide von Metallen der Übergangsgruppen (im Periodensystem). Auf der Grundlage dieses Vorgangs nahmen Anbar et al. (2007) an, dass eine Anreicherung der Atmosphäre mit Sauerstoff zu einer erhöhten Löslichkeit von Molybdän- bzw. Rhenium-Sulfiden geführt haben sollte. Demnach soll der Nachweis erhöhter Konzentrationen an Molybdat (MoO42-) in Gesteinen ein Hinweis auf eine Anreicherung der Atmosphäre mit Sauerstoff in früherer geologischer Zeit sein. Die Ergebnisse ihrer geochemischen Studien an Gesteinen des Mount McRae Shale in der westaustralischen Hamersley-Senke interpretierten Anbar et al. dahingehend, dass es schon vor 2,5 Milliarden Jahren Sauerstoff in der Erdatmosphäre gegeben haben muss, wenn auch in geringer Konzentration. Deshalb war in der entsprechenden Publikation die Rede von „einem Hauch Sauerstoff“ vor dem eigentlichen GOE. Noch früher datierten Koehler et al. den schleichenden Beginn des GOE. Ihren Ausführungen zufolge (s. u. „Isotopensignaturen“) soll es bereits vor 2,66 Milliarden radiometrischen Jahren Sauerstoff in der Erdatmosphäre gegeben haben. Die außerordentlich diffusen und widersprüchlichen Angaben über das Einsetzen des GOE werfen die Frage auf, ob es jemals wirklich einen klaren Übergang von einer anoxischen zu einer oxischen Atmosphäre gegeben hat.
Isotopensignaturen
Als weiteres Indiz für den GOE werden verschiedene Isotopensignaturen angeführt. Mit dem Begriff „Isotopensignaturen“ werden Mengenverhältnisse von Isotopen* bestimmter Elemente bezeichnet, die in den verschiedenen Gesteinen durchaus variieren können.
Schwefelisotope
Besondere Relevanz im Zusammenhang mit dem GOE haben Messungen von Isotopenverhältnissen des Elements Schwefel erhalten. Gewöhnlich ist von den vier stabilen Isotopen des Schwefels 32S das am häufigsten vorkommende (95,02%), gefolgt von 34S (4,21%), 33S (0,75%) und 36S (0,02%). Bei vielen physikalischen Prozessen oder biologischen Vorgängen beobachtet man eine Auftrennung der verschiedenen Isotopen eines chemischen Elements, z. B. bei der Verstoffwechslung durch Bakterien oder bei Diffusionsvorgängen. Die Beobachtung, dass die Trennung der Isotopen voneinander (Fraktionierung) proportional zu ihrer Massendifferenz geschieht, wird als massenabhängige Isotopenfraktionierung bezeichnet. Durch solche Vorgänge ändert sich z. B. das Verhältnis von 34S/32S etwa halb so schnell wie das Verhältnis 36S/32S, da die Massendifferenz zwischen 34S und 32S etwa halb so groß ist wie diejenige von 36S und 32S. Isotopenfraktionierungen, die mit signifikanten Abweichungen von dieser Gesetzmäßigkeit einhergehen, gelten dementsprechend als massenunabhängige Fraktionierungen. Die diesen Beobachtungen zugrundeliegenden Mechanismen sind aber bisher nicht aufgeklärt (Dauphas & Schauble 2016).
Das Bild kann online nicht zur Verfügung gestellt werden.
Abb. 3: Auftragung der Isotopenfraktionierung von 33S (in ‰) gegen das ermittelte Alter der entsprechenden Proben (in Millionen radiometrischen Jahren) aus Thiemens (2013). Die roten Punkte repräsentieren Proben, in denen eine massenunabhängige Fraktionierung des Schwefelisotops nachgewiesen wurde, während die blauen Punkte Proben mit massenabhängigen Fraktionierungen darstellen.
Bei Untersuchungen an präkambrischen Gesteinen wurden massenunabhängige Fraktionierungen von Schwefelisotopen nachgewiesen (Farquhar et al. 2000a), die z. T. signifikant von den Standardwerten abweichen (s. Abb. 3).
Zudem ist bei Laborversuchen gezeigt worden, dass die Einwirkung von UV-Licht auf Schwefeldioxid ebenfalls ähnliche massenunabhängige Isotopenverteilungen des Schwefels bewirkt (Farquhar et al. 2000b). Diese Befunde wurden als Indizien dafür gewertet, dass es in der Zeit vor ca. 2,3 Milliarden radiometrischen Jahren eine sauerstofffreie Erdatmosphäre gegeben haben soll. Denn bei Abwesenheit von Sauerstoff fehlt auch die Ozonschicht, sodass aus Vulkaneruptionen stammendes Schwefeldioxid der UV-Strahlung ausgesetzt gewesen sein müsste. Das Verschwinden der massenunabhängigen Isotopenfraktionierung des Schwefels wird daher als Beleg für das Einsetzen der globalen Anreicherung der Erdatmosphäre mit Sauerstoff interpretiert.
Auch Luo et al. (2016) verwendeten die Fraktionierung von Schwefelisotopen, um Rückschlüsse über den GOE anhand von Untersuchungen an Pyritgestein aus einer Sedimentabfolge der Transvaal Group in Südafrika zu ziehen. Sie ordneten das Ereignis ebenfalls auf ca. 2,3 Milliarden radiometrische Jahre vor unserer Zeit ein. Zudem gaben sie noch an, dass der Übergang zu einer mit Sauerstoff angereicherten Atmosphäre sich in nur 1-10 Millionen Jahren ereignet haben soll – eine sehr kurze Zeit (bei Voraussetzung von Evolution) für die tiefgreifende Umstellung des Stoffwechsels der großen Mehrheit der Mikroorganismen auf den chemisch ausgesprochen reaktiven Sauerstoff.
Zu einem völlig anderen Ergebnis kamen dagegen Blättler et al. (2018). Sie untersuchten Evaporitgestein aus einer Tiefe von 2100-2900 m aus der Onega-Senke in Russland und analysierten ebenfalls schwefelhaltige Mineralien hinsichtlich der Isotopenfraktionierung des Schwefels. Ihren Ausführungen zufolge handelte es sich bei dem GOE um eine „langwierige Reorganisation globalen Redox-Budgets“, die Hunderte Millionen von Jahren in Anspruch nahm.
Die Verwendung der Analyse von Schwefelisotopenmustern zur Beurteilung der Zusammensetzung der Atmosphäre im Proterozoikum hat sich ungeachtet widersprüchlicher Schlussfolgerungen etabliert. Allerdings ist bis heute unklar, was die eigentliche physikalisch-chemische Ursache der massenunabhängigen Isotopenfraktionierung ist. Auch ist es fraglich, ob durch die Laborexperimente von Farquhar et al. die komplexen Bedingungen der Atmosphäre realistisch simuliert worden sind.
Auswertungen von Daten zu Isotopenfraktionierungen führen je nach untersuchtem Element zu sehr unterschiedlichen Schlussfolgerungen hinsichtlich des GOE.
Weitere Analysen anhand von Isotopenfraktionierungen
Als eine weitere Möglichkeit, Informationen über die Zustände der Atmosphäre und Biosphäre zu erhalten, werden weitere gegenüber der Oxidation empfindliche chemische Elemente verwendet. Koehler et al. (2018) werteten Isotopenverhältnisse des Stickstoffs und des Selens von Gesteinen der Jeerinah-Formation in Australien aus. Bestimmte Isotopenmuster des Stickstoffs werden als möglicher Indikator für Nitrat-verstoffwechselnde (denitrifizierende) Mikroorganismen aufgefasst. Interessanterweise kamen Koehler et al. auf der Grundlage ihrer Befunde zu dem Ergebnis, dass es schon bereits vor 3,2 Milliarden Jahren einen Stickstoffkreislauf gegeben haben muss. Ein Stickstoff-Kreislauf kann jedoch nur bei Vorhandensein von Sauerstoff etabliert sein. Demnach müsste es schon vor 3,2 Milliarden Jahren zumindest in geringen Konzentrationen Sauerstoff in der Erdatmosphäre gegeben haben – also schon 800 Millionen Jahre vor dem eigentlichen GOE. Die zeitlichen Verläufe der verschiedenen Modelle des GOE unterscheiden sich um viele Hunderte von Millionen von Jahren.
Auch über den Verlauf des GOE sind sich die Forscher uneinig: einige schlussfolgern, dass die Anreicherung der Atmosphäre sich schnell ereignete, während andere der Auffassung sind, dass sich diese Umstellung sehr langsam vollzog. Es zeigt sich also insgesamt, dass die Interpretationen der geologischen Daten zur Rekonstruktion des GOE sehr vage und widersprüchlich sind.
Glossar
Ad-hoc-Annahme: Annahme, für die es keine empirischen Indizien gibt. aerob: Sauerstoff verstoffwechselnd. anaerob: unter Ausschluss von Sauerstoff. anoxisch: frei von Sauerstoff. Archaikum: erdgeschichtliche Epoche von vor 4,0 Milliarden bis 2,5 Milliarden radiometrischen Jahren. Hadaikum: erdgeschichtliche Epoche von vor 4,6 Milliarden bis 4,0 Milliarden radiometrischen Jahren. Isotop: Ein chemisches Element ist durch die Anzahl an Protonen im Atomkern definiert. Atome desselben Elements können sich aber hinsichtlich der Anzahl an Neutronen unterscheiden. In diesem Fall spricht man von verschiedenen Isotopen eines chemischen Elements. Oxidantien: Chemische Verbindungen, die anderen Stoffen Elektronen entziehen. oxidierend: chemische Verbindungen mit der Neigung, Elektronen aufzunehmen, heißen „oxidierende“ Verbindungen. In einer „oxidierenden Atmosphäre“ dominieren solche Stoffe. oxygen: Sauerstoff erzeugend. Oxygenierung: Anreicherung mit Sauerstoff. Phanerozoikum: erdgeschichtliche Epoche von vor 541 Millionen radiometrischen Jahren bis heute. photoautotroph: Bezeichnung für die Lebensweise von Organismen, die ihre Energie aus Licht beziehen können. Als Elektronenquelle dient in diesem Fall Wasser. photoferrotroph: Bezeichnung für die Lebensweise von Organismen, die ihre Energie aus Licht beziehen können und als Elektronenquelle Eisen(II)ionen verwenden. Proterozoikum: erdgeschichtliche Epoche von vor 2500 Millionen bis vor 541 Millionen radiometrischen Jahren. reduzierend: chemische Verbindungen mit der Neigung, Elektronen abzugeben, heißen „reduzierende“ Verbindungen. In einer „reduzierenden Atmosphäre“ dominieren solche Stoffe.
Paläobiologische Indizien
Mikrofossilien
Das Bild kann online nicht zur Verfügung gestellt werden.
Abb. 4: A und D Ramsaysphaera ramses, fossil, datiert auf 3,4 Milliarden rad. Jahre; B und C Candida tropicalis, rezent (nach Pflug 1978); E-J Mikrofossilien aus dem frühen Archaikum (Apex Chert von Westaustralien, nach Schopf 1993)
Gewöhnlich werden im Rahmen der Evolutionslehre Veränderungen von Lebewesen, die in einer weit zurückliegenden Vergangenheit liegen sollen, durch Interpretation von Fossilfunden rekonstruiert. Dies hat sich im Hinblick auf die Frage nach den ältesten fossil dokumentierten Organismen, die Sauerstoff durch Photosynthese erzeugten, als schwierig erwiesen. Wann die ersten Mikroorganismen, die oxygene* Photosynthese betrieben, entstanden sein sollen, war lange Zeit umstritten. Butterfield (2000) beschrieb fossile Rotalgen aus der Hunting-Formation auf Somerset Island in Canada, die auf ein Alter von 1,2 Milliarden radiometrische Jahre datiert worden ist. Allerdings gibt es eine ganze Reihe an Arbeiten, die darüber Auskunft geben, dass es schon viel früher oxygene Photosynthese betreibende Organismen gab. Der Nachweis fossiler, fadenförmiger Kolonien von Cyanobakterien im Apex Chert in Westaustralien (Schopf 1993, Abb. 4), die auf über 3,4 Milliarden radiometrische Jahre datiert wurden, und Hinweise auf photosynthetisch aktive Mikroorganismen vor 3,1 Milliarden radiometrischen Jahren in verschiedenen Formationen in Südafrika (Tice & Lowe 2004) ergeben ein anderes Bild. In einer Übersichtsarbeit zu Mikrofossilien aus dem Archaikum* legte Schopf dar, dass 48 Nachweise von Stromatolithen* hauptsächlich photoautotropher* Mikroorganismen dokumentiert wurden (Schopf 2006). Diesen Daten zufolge hätte es schon vor über 3,4 Milliarden Jahren sauerstofferzeugende Organismen geben müssen, womit der Beginn des GOE um eine Milliarde Jahre früher eingesetzt hätte als nach der konventionellen Darstellung.
Dies wäre jedoch problematisch für die Evolutionstheorie, da dann erklärt werden müsste, weshalb schon die allerersten Organismen oxygene Photosynthese betreiben konnten. Für die Evolution dieser hochkomplexen biochemischen Einrichtungen wäre dann nach evolutionären Maßstäben kaum Zeit vorhanden gewesen, da allgemein angenommen wird, dass der Beginn des Lebens sich vor ca. 3,8–4,1 Milliarden radiometrischen Jahren vor unserer Zeit ereignet haben soll. Es überrascht daher nicht, dass Homann-Marriott & Blankenship (2011) die Befunde von Schopf grundsätzlich in Frage stellen: „Es wurden jedoch kürzlich Zweifel bezüglich der Ungestörtheit der Geologie des Fundortes und der Eignung der Umweltbedingungen für mikrobielles Leben sowie der Fossilien selbst geäußert.“
Hierzu sei angemerkt, dass bei Mikrofossilien solche Zweifel prinzipiell immer möglich sind. Ob der Fundort für mikrobielles Leben geeignet und durchweg ungestört war, kann angesichts des bisher noch geringen Wissens über die Lebensweise von Mikroorganismen in Gesteinen nicht sicher beurteilt werden. Auch der Interpretationsspielraum bezüglich der biochemischen Ausstattung der fossil nachgewiesenen Mikroorganismen scheint gemäß Schopf selbst recht weit zu sein, denn einerseits heißt es in seiner Veröffentlichung von 2006: „Daher wurde allgemein angenommen, dass das Vorhandensein dieser morphologisch mutmaßlich Cyanobakterien-ähnlichen Fossilien darauf hinweist, dass hochgradig entwickelte Prozesse der mikrobiellen Biochemie wie die oxygene Photosynthese und die aerobe Zellatmung bereits vor 3,5 Milliarden Jahren entwickelt waren.“ Andererseits heißt es etwas weiter im Text: „Es ist denkbar, dass die äußerliche Ähnlichkeit der Apex-Mikroorganismen [Fossilien] zu jüngeren sauerstoffproduzierenden Cyanobakterien signifikante Unterschiede der inneren Biochemie maskiert.“
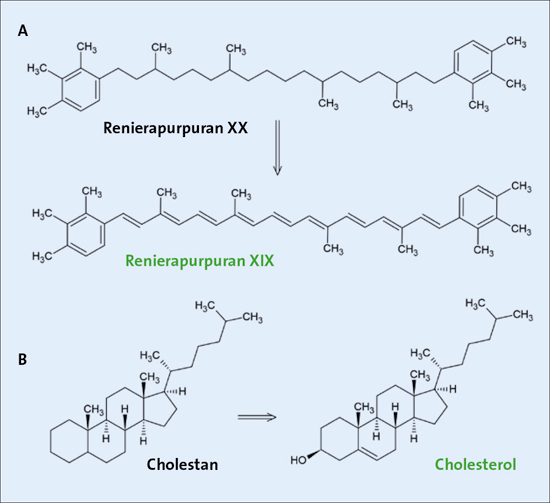
Abb. 5: Strukturformeln zweier Moleküle, die als „molekulare Marker“ (Bezeichnung in schwarzer Schrift) gelten und ihre vermutlichen Vorstufen aus Organismen (Bezeichnung in grüner Schrift). Das Renierapurpurin XIX ist ein Bestandteil von Cyanobakterien, während Cholesterol ein typischer Baustein der Zellwand von Eukaryoten ist.
Hier wird also eine naheliegende Interpretation (äußere morphologische Ähnlichkeit der Fossilien zu oxygenen Mikroorganismen) durch vage Spekulationen über die nicht nachweisbare innere Biochemie der mutmaßlichen früheren Organismen in Frage gestellt, was nicht gerade überzeugend ist. In späteren Arbeiten bekräftigte Schopf (2017; 2018) das konventionelle Modell des GOE, allerdings unter Verweis auf Isotopendaten. Obwohl, wie bereits erwähnt, anhand der äußerlichen Merkmale der Mikrofossilien keine sichere Aussage über die innere Biochemie der früheren Mikroorganismen gemacht werden kann, sprach sich Schopf in seinen letzten Arbeiten dafür aus, dass die photosynthetisch aktiven Bakterien keinen Sauerstoff produzierten, sondern nicht-oxygene Photosynthese betrieben. Die oxygene Photosynthese sei dann in einem späteren Stadium der mikrobiellen Evolution entstanden.
Insgesamt sieht die Datenlage bezüglich fossilisierter Mikroorganismen, die mittels Photosynthese Sauerstoff erzeugen konnten, widersprüchlich und unklar aus. Ein Umschwung von einer nicht oxygenen zu einer Sauerstoff-produzierenden mikrobiellen Welt ist jedenfalls anhand der Mikrofossilien nicht nachweisbar. Dass Schopf als einer der größten Experten auf dem Gebiet der Mikrofossilien sich von der früheren Interpretation seiner Ergebnisse abwandte, um das Standardmodell zu befürworten, scheint nicht die Folge einer zwingenden Indizienlast zu sein.
Molekulare Marker
Nicht wenige Forscher haben konstatiert, dass der Fossilbericht alles andere als reich ist an fossilen Resten von Mikroorganismen, die zur Zeit des GOE lebten (Brocks et al. 1999; Gueneli et al. 2018; Cui et al 2020). Aus diesem Grund ist nach alternativen Hinweisen auf frühes Leben gesucht worden. Als solche werden aus Gesteinen entnommene organische Moleküle gewertet, die als „Biomarker“ bezeichnet werden. Cui et al. (2020) beschrieben Gemische aus Carotinoiden, die sie als fossilisierte molekulare Spuren von Cyanobakterien interpretieren. Allerdings handelt es sich dabei nicht um genau diejenigen Moleküle, die auch in heutigen Cyanobakterien vorgefunden werden, sondern um durch unbekannte chemische Reaktionsfolgen erhaltene Derivate (s. Abb. 5). Um diese Annahme zu plausibilisieren, werden die unbekannten Reaktionsfolgen durch Simulationen im Labor nachgestellt. Es ist allergings fraglich, ob chemische Reaktionsfolgen im Labor geeignet sind, um geochemische Vorgänge zu simulieren.
Sichere Aussagen über die Art und Beschaffenheit früher Mikroorganismen sind weder anhand von Mikrofossilien noch Biomarker-Molekülen möglich.
Ein weiteres grundsätzliches Problem bei dem Nachweis von Biomarkern sind Kontaminationen. Brocks & Summons (1999) beschreiben dieses Problem im Zusammenhang mit einem überraschenden Nachweis von Steranen (z. B. Cholestan, s. Abb. 5) aus 2,7 Milliarden radiometrische Jahre altem Gestein wie folgt: „Obwohl diese Gesteine für Verhältnisse des Archaikums gut erhalten sind, sind sie dennoch sehr alt und enthalten daher nur geringe Mengen an extrahierbaren Kohlenwasserstoffen. Daher sind sie besonders anfällig für moderne Verunreinigungen durch Erdölprodukte während des Bohrens, der Lagerung und der Analyse.“ Sterane gelten als molekulare Spuren von eukaryoten Organismen und würden den konventionellen Vorstellungen der Evolution früher Organismen völlig widersprechen. Diese sensationellen Befunde wurden jedoch später aufgrund von angeblichen Kontaminationen der Proben verworfen (Gold et al. 2017). Insgesamt erscheint es also fraglich, ob es sich bei den als „molekulare Fossilien“ bezeichneten organischen Molekülen um sichere Hinweise auf frühere Mikroorganismen handelt.
Fazit
Die Frage nach dem Ursprung des Sauerstoffs in der Atmosphäre hat in den vergangenen vier Jahrzehnten überraschend viel Aufmerksamkeit erfahren. Ungeachtet des großen Forschungsaufwandes gibt es in vielen Bereichen nach wie vor nur sehr vage Ergebnisse und widersprüchliche Schlussfolgerungen bezüglich der Zeit des Einsetzens des GOE und seines Verlaufs. Der Konsens, dass das GOE vor ca. 2,4 Milliarden radiometrischen Jahren eingesetzt haben soll, beruht auf einer unzuverlässigen Datenbasis und vielen spekulativen Annahmen. Probleme wie ein durch den starken Treibhauseffekt des Methans resultierendes Backofen-Klima oder durch Anreicherung von Sauerstoff resultierende dauerhafte globale Vereisungen oder das Fehlen einer schützenden Ozon-Schicht vor dem Einsetzen des GOE werden in der Literatur bisher nicht in gebührender Weise behandelt. Die Frage, wie es zur Feinabstimmung der vielen Parameter wie Intensität der Sonneneinstrahlung, Zusammensetzung der Atmosphäre und Beschaffenheit der Biosphäre kam, so dass die Erde ein lebensfreundlicher Planet wurde, wird gar nicht thematisiert. Jedenfalls scheint die passende Einstellung dieser Parameter durch rein natürliche Vorgänge extrem unwahrscheinlich zu sein. Insgesamt sind die geologischen und paläobiologischen Indizien zum postulierten GOE unklar und inkonsistent, sodass Hypothesen einer frühen Evolution von Mikroorganismen damit nicht gut untermauert werden können. Noch weit schwieriger ist es jedoch, die Umstellung des Stoffwechsels von anaerob auf aerob durch Evolution zu erklären. Dieser Fragestellung wird eine weitere Folge dieser Artikelserie gewidmet sein.
1 „The continuing puzzle of the great oxydation event.“
2 https://www.lubw.baden-wuerttemberg.de/luft/zusammensetzung-der-luft, abgerufen am 10. 3. 2021.
3 https://www.dw.com/de/methan-der-b%C3%B6se-zwillingsbruder-von-co2/a-49208882, abgerufen am 22. 2. 21 mit Verweis auf Nisbet et al. (2019).
A Whiff of Oxygen Before the Great Oxidation Event? Science 317, 1903–1906.
Two-billion-year-old evaporites capture earth’s great oxidation. Science 360, 320–323.
Bangiomorpha pubescens n. gen., n. sp.: implications for the evolution of sex, multicellularity, and the Mesoproterozoic/Neoproterozoic radiation of eukaryotes. Paleobiology 26, 386–404.
Archean molecular fossils and the early rise of eukaryotes. Science 285, 1033–1036.
Paleological significance of the banded Iron-Formation. Economic Geology 68, 1133–1143.
Niche expansion for phototrophic sulfur bacteria at the Proterozoic-Phanerozoic transition. PNAS 117, 17599–17606.
Mass-independent effects, and isotopic anomalies. Annu. Rev. Earth. Planet. Sci. 44, 709–783.
a.) Atmospheric Influence of earths earliest sulfur cycle. Science 289, 756–758; b.) Evidence of atmospheric sulphur in the martian regolith from sulphur isotopes in meteorites. Nature 404, 50–52.
Iron formation in neoarchaean deltaic successions and the microbially mediated deposition of transgressive systems tracts. J. Sediment. Res. 76, 1057–1066.
Banded Iron-Formations through much of Precambrian time. J. Geol. 89, 169–183.
1.1-billion-year-old porphyrins establish a marine ecosystem dominated by bacterial primary producers. PNAS 115, E6978–E6986.
Timing and tempo of the great oxidation event. PNAS 114, 1811–1816.
Evolution of photosynthesis. Annu. Rev. Plant. Biol. 62, 515–548.
The timetable of evolution. Sci. Adv. 3: e1603076.
Transient surface ocean oxygenation recorded in the 2,66 Ga Jeerinah Formation, Australia. PNAS 115, E8323.
Rapid oxygenation of Earth’s atmosphere 2.33 billion years ago. Sci. Adv. 2: e1600134.
A production of amino acids under possible primitive earth conditions. Science 117, 528–529.
Organic compound synthesis on the primitive Earth. Science 130, 245–251.
Very strong atmospheric methane growth in the 4 years 2014–2017: Implications for the Paris agreement. Global Biogeochemical Cycles 33, 318–342.
Die Entstehung des Lebens auf der Erde. Leipzig.
Yeast-like microfossils detected in oldest sediments of the earth. Naturwissenschaften 65, 611–615.
Microfossils of the early archean apex chert: new evidence of the antiquity of life. Science 260, 640–646.
Fossil evidence of archaean life. Phil. Trans. R. Soc. B 361, 869–885.
An anaerobic 3400 Ma shallow –water microbial consortium: presumptive evidence of earth’s paleoarchean anoxic atmosphere. Precambrian Research 299, 309–318.
Sims analyses of the oldest known assemblage of microfossils document their taxon-correlated carbon isotope compositions. PNAS 115, 53–58.
The continuing puzzle of the great oxidation event. Curr. Biol. 19, 567–574.
Introduction to Chemistry and Applications in Nature of Mass Independent Isotope Effects Special Feature. PNAS 110, 17631–17637.
Themen | Kurzbeiträge | Streiflichter
Studiengemeinschaft WORT und WISSEN e.V.
Letzte Änderung: 7/2/21
Webmaster