Themen • Kurzbeiträge • Streiflichter
Vogelmerkmale bei Dinosauriern
Vorläuferstadien oder Konvergenzen?
von Reinhard Junker
Studium Integrale Journal
27. Jahrgang / Heft 2 - Oktober 2020
Seite 68 - 77
Zusammenfassung: „Vögel sind Dinosaurier“ – das ist heute die kaum angefochtene Lehrmeinung zum Ursprung der Vögel. Die wichtigste Begründung: Die meisten vogeltypischen Merkmale seien schrittweise bei Dinosauriern entstanden. Eine genauere Analyse der fossilen Überlieferung offenbart jedoch ein anderes Bild.
• • • • • • • •
Einführung
„Vögel sind heute lebende Dinosaurier“ (Erickson et al. 2017). „In Wahrheit sind Vögel Dinosaurier“ (Brusatte 2017a, 531). Man mag diese Gleichsetzung von Vögeln und Dinosauriern sonderbar oder gar abwegig finden, aber in den letzten Jahrzehnten hat sich die mutmaßliche Abstammung der Vögel von Dinosauriern als einer der populärsten evolutionären Übergänge in der Fachwelt fast unangefochten etabliert.2
Mit einem Stern* versehene Begriffe werden im Glossar erklärt.
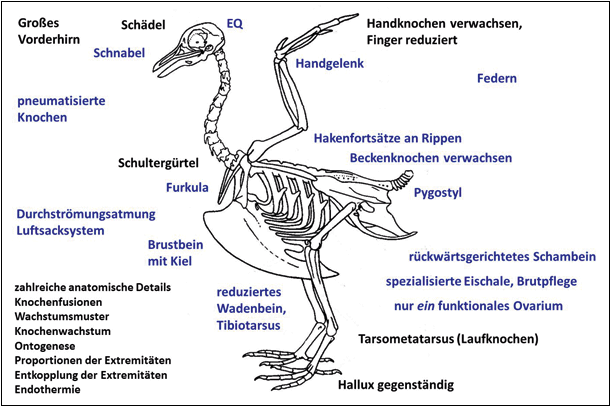
Abb. 1: Überblick über typische Vogelmerkmale (nicht vollständig): In der Analyse (Junker 2019) wurden die blau gesetzten Merkmale berücksichtigt. In diesem Beitrag wird eine repräsentative Auswahl vorgestellt und diskutiert. (Skelett: CC BY 2.5)
In den letzten Jahren wurde eine Reihe von Studien veröffentlicht, die einen schrittweisen Übergang von Dinosauriern zu Vögeln dokumentieren sollen (vgl. Abb. 2). Die Merkmale, die charakteristisch für Vögel sind (vgl. Abb. 1), seien im Wesentlichen in unterschiedlicher Häufigkeit bereits bei verschiedenen Dinosaurier-Gattungen aus mehreren Familien der Theropoden ausgebildet gewesen (vgl. Abb. 3). Man könne daher heute keine Grenze mehr zwischen Dinosauriern und Vögeln ziehen (Brusatte 2017a, 553). Zu den vogeltypischen Merkmalen bei Dinosauriern gehören allen voran Federn oder federartige Körperanhänge4, das Gabelbein (Furkula), ein großes Brustbein, verknöcherte Brustbeinplatten, ein vergrößerter Encephalisations-Quotient, lange Arme, der Bau des Handgelenks5, eine dreifingrige Hand mit langem zweitem Finger6, hohle Knochen, das Luftsacksystem, hakenförmige, nach hinten gerichtete Fortsätze an den vorderen Rippen7, ein nach hinten gerichtetes Schambein, zweibeinige Fortbewegung, lange Hinterbeine mit dreizehigem Fuß, ein reduziertes Wadenbein, verschmolzene Schwanzwirbel, Brutpflege, eine deutlich verringerte Körpergröße und andere (Padian & Chiappe 1998, 44; Organ et al. 2007; Xu 2006; Xu et al. 2014; vgl. Abb. 1).
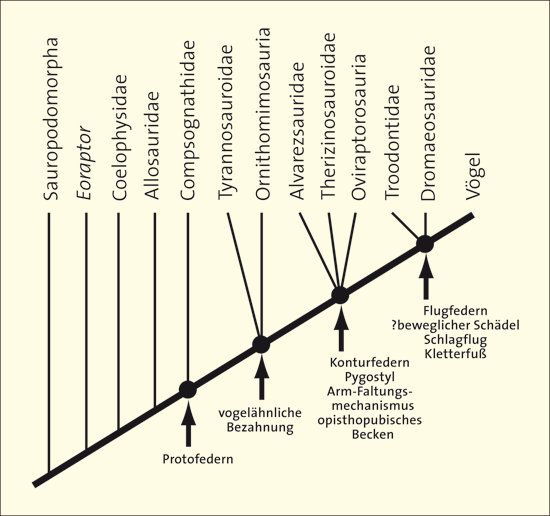
Abb. 2: Beispiel einer Darstellung eines schrittweisen Erwerbs von Vogelmerkmalen bei Theropoden-Dinosauriern. (Nach Xu 2006)
Umgekehrt besitzen viele Vögel aus dem Oberjura (allen voran der berühmte „Urvogel“ Archaeopteryx) und aus der Unterkreide Merkmale, die für heutige Vögel untypisch sind, jedoch bei vielen Theropoden-Gattungen ausgebildet sind. Dazu gehört der verbreitete Besitz von Zähnen, eine lange Schwanzwirbelsäule (bei Archaeopteryx und Jeholornis), das Fehlen eines Brustbeinkiels, relativ wenige Knochenverwachsungen und weitere Merkmale.
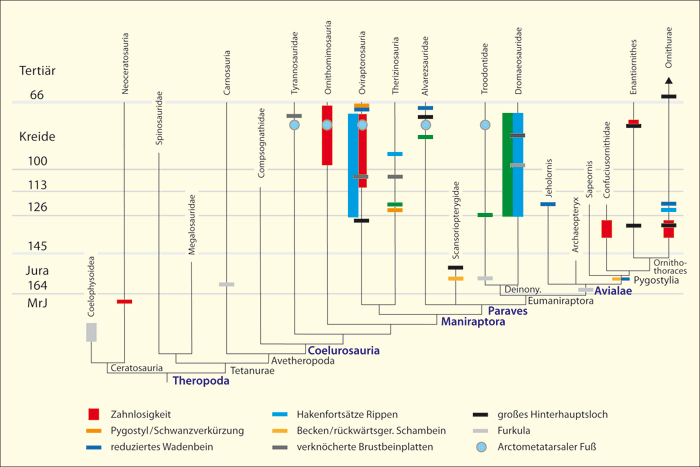
Abb. 3: Zeitskaliertes Cladogramm der Theropoden-Dinosaurier-Gruppen und Vögel und Zusammenschau der konvergenten Verteilung einiger vogeltypischer Merkmale. Die stammesgeschichtlichen Rekonstruktionen verschiedener Bearbeiter ergeben häufig unterschiedliche Anordnungen einzelner Gruppen. An der vielfachen Konvergenz ändern die verschiedenen Cladogramm-Varianten jedoch nichts. Die Merkmalsverteilungen sind vielfach so unsystematisch, dass die Annahme zahlreicher Konvergenzen in jedem Fall unvermeidbar ist. MrJ: Millionen radiometrische Jahre. Arctometatarsaler Fuß ist eine spezielle Form des Fußes. (Nach zahlreichen Quellen zusammengestellt, vgl. Junker 2019)
So gesehen scheint es gut begründet, dass es einen fast durchgehend graduellen Übergang von theropoden Dinosauriern zu den Vögeln gibt. Diese Einschätzung wird dadurch unterstützt, dass innerhalb der einzelnen relevanten Merkmale auch noch unterschiedliche Ausprägungen vorkommen, beispielsweise ein unterschiedliches Ausmaß der Reduktion von Zähnen oder unterschiedlich ausgeprägte Pygostyle* (s. u.).
Doch dieses Bild ist unvollständig und dadurch mehr oder weniger irreführend und zwar aus folgenden Gründen:
1. Viele Vogelmerkmale bei Dinosauriern treten mehrfach unabhängig (konvergent*) auf, also nicht nur in einer einzigen, sondern in verschiedenen Linien ohne gemeinsamen Vorfahren. Dementsprechend sind die einzelnen Vogelmerkmale teilweise auf verschiedenen Ästen der Theropoden verteilt (Abb. 3).
2. Die stratigraphischen* Positionen der Dinosaurier-Gattungen, die eine unterschiedliche Anzahl vogeltypischer Merkmale besitzen, entsprechen vielfach nicht den evolutionär anzunehmenden zeitlichen Abfolgen (vgl. Abb. 3).
3. Entsprechendes gilt für Dinosauriermerkmale und ihre Abfolgen bei den Oberjura- und Unterkreidevögeln.
4. Manche Gattungen, die als Vogelvorläufer interpretiert werden, könnten sekundär flugunfähige Vögel sein. Deren Merkmale würden in diesem Falle als Belege für einen schrittweisen Übergang von Dinosauriern zu Vögeln ausscheiden.
5. Bei manchen Merkmalen gibt es mehr oder weniger markante Sprünge (wo schrittweiser Umbau funktionell problematisch wäre).
Diese Situation äußert sich z. B. darin, dass keine konkrete Gattung als (letzter) Vorfahr der Vögel angegeben werden kann: „Obwohl praktisch alle neueren Analysen die Dromaeosauridae oder Troodontidae … als Schwestergruppe der Vögel ausweisen, ist keiner von beiden wirklich der Vorfahre, und daher können bekannte Formen wie Deinonychus oder Troodon nur als Modelle für den wahren Vogelvorfahren dienen“ (Witmer 2002, 168).
Kompakt
In den letzten Jahrzehnten hat sich die mutmaßliche Abstammung der Vögel von Dinosauriern als einer der populärsten evolutionären Übergänge etabliert. Als Begründung wird dafür genannt, dass ein Großteil von Vogelmerkmalen bei verschiedenen Theropoden-Dinosauriern vertreten sei. Es könne an Fossilien nachvollzogen werden, dass Vogelmerkmale schrittweise aufgetreten seien.
Eine detaillierte Analyse zeigt jedoch, dass zahlreiche vogeltypische Merkmale, die bei Theropoden-Dinosauriern vorkommen, evolutionstheoretisch gedeutet mehrfach unabhängig entstanden sein müssen. Gemäß aktueller Phylogenien* und nach dem Fossilbefund waren einige dieser Merkmale an der Basis der jeweiligen Gruppen gar nicht ausgeprägt. Teilweise werden entsprechend je nach zugrundgelegtem Merkmal verschiedene Verwandtschaftsverhältnisse nahegelegt, was mit vernetzten, mosaikartigen Merkmalsbeziehungen einhergeht und einem einfachen Verzweigungsschema („Stammbaum“) widerspricht. Diese häufig mosaikartige Verteilung begünstigt auch die umstrittene Deutung, dass manche Gattungen, die als Vogelvorläufer interpretiert werden, in Wirklichkeit sekundär flugunfähige Vögel sein könnten. Deren Merkmale würden in diesem Falle als Belege für einen schrittweisen Übergang von Dinosauriern zu Vögeln ausscheiden.
Aufgrund dieser Befunde wird die Behauptung eines schrittweisen evolutionären „Aufbaus“ des Vogelbauplans anhand des Fossilbefundes stark in Frage gestellt und sie wird bei zahlreichen Merkmalen durch den Fossilbefund kaum unterstützt. Die meisten Theropoden-Dinosaurier-Gattungen, die vogelartige Merkmale besitzen, sind zudem geologisch (z. T. deutlich) jünger als die geologisch ältesten Vögel.
Vogelmerkmale bei Theropoden: Vorläufer oder Konvergenzen?
Im Folgenden soll an vier wichtigen vogeltypischen Merkmalen bei Gattungen, die zu Dinosauriern gestellt werden, untersucht werden, was über ihre Verteilung bekannt ist. Hintergrund ist die eingangs genannte Behauptung, die meisten Vogelmerkmale seien Schritt für Schritt in Dinosaurierlinien entstanden. Lässt sich das anhand der Merkmalsverteilung verifizieren? Und passen die stratigraphischen Positionen derjenigen Gattungen, die als Vorläufer interpretiert werden, zu den stammesgeschichtlichen Rekonstruktionen? Betrachtet werden Federn, Furkula, Hornschnabel und Pygostyl. Diese Merkmale sowie elf weitere werden in Junker (2019) ausführlicher diskutiert. Hier erfolgt eine kompakte Darstellung von vier repräsentativen Merkmalen.
Glossar
abgeleitet: im Stammbaum weit oben stehend, „höherentwickelt“. Cladogramm: Gabelig-verzweigte Darstellung von Ähnlichkeitsbeziehungen. Enantiornithes: Nur in Sedimenten der Kreide fossil überlieferte Vogelgruppe, deren besonderes Kennzeichen die Gelenkverbindung zwischen Schulterblatt und Rabenbein ist. Am Schulterblatt befindet sich eine Gelenkpfanne und am Rabenbein ein Gelenkzapfen, anders herum als bei anderen Vogelgruppen. Furkula: Gabelbein; bei Vögeln und einigen Dinosauriern aus den zwei Schlüsselbeinen verschmolzenes, gabelig geformtes Knochenelement. Gegenvögel: → Enantiornithes. Grundtyp: Arten, die direkt oder indirekt kreuzbar sind, gehören zu einem Grundtyp. Konvergenz: gleichartige Ausprägung von Strukturen abstammungsmäßig nicht verwandter Organismen, die evolutionstheoretisch als unabhängig entstanden interpretiert werden. Ornithuren: „Vogelschwänze“; Vogel mit Fächerschwanz, zu denen auch die heutigen Vögel gehören; ab der Unterkreide fossil überliefert. Paraves: Vögel und ihre evolutionstheoretisch nächstverwandten Gruppen: Üblicherweise werden die Deinonychosaurier (Troodontidae und Dromaeosauridae) dazu gerechnet (vgl. Abb. 3). Phylogenie: Evolutionstheoretisch rekonstruierte Abstammungsabfolge, ermittelt durch cladistische Analysen. plesiomorph: Bezeichnung für ein Merkmal oder einen Merkmalszustand, das bzw. der evolutionstheoretisch als primitiv bzw. ursprünglich gewertet wird. Pygostyl: Durch die Verschmelzung mehrerer Wirbel gebildeter Knochen am Ende der Wirbelsäule von Vögeln.Pygostylia: Vögel, die ein → Pygostyl besitzen. Stratigraphie: Schichtbeschreibung, Abfolge von geologischen Sedimentschichten. Taxon (pl. Taxa): Einheit der Klassifikation, seien es Arten, Gattungen, Familien oder andere Gruppen von Lebewesen, die gewöhnlich als abstammungsbedingte Einheiten angesehen werden.
Federtypen und Flugfähigkeit
Zitate zu schrittweisem Erwerb von Vogelmerkmalen finden sich im Online-Zusatzmaterial.
Bei einer großen Anzahl von Theropodengattungen, die systematisch in die Nähe der Vögel gestellt werden, wurden haarartige oder federähnliche Anhänge oder auch echte flächige Federn fossil nachgewiesen. Dabei bestehen deutliche Unterschiede zwischen haarartigen, flaumigen (ggf. verzweigten) oder borstenartigen Anhängen einerseits und flächigen, flugtauglichen Federn andererseits (Überblick und Quellenbelege: Junker 2017). Nach Brusatte (2017b, 792) war die Entwicklung der Flugfähigkeit „chaotisch“; verschiedene Dinosaurier hätten mit verschiedenen luftgestützten Verhaltensweisen und verschiedenen Feder-Anordnungen „experimentiert“, bis schließlich nur die modernen Vögel überlebt haben.9 Abgesehen davon, dass der Begriff „experimentieren“ evolutionstheoretisch höchst problematisch ist, geht aus diesem Zitat hervor, dass von einer linearen, schrittweisen Entwicklung nicht die Rede sein kann. Vielmehr sind (in evolutionstheoretischer Lesart) in verschiedenen Linien unterschiedliche Ausprägungen von Flugfähigkeit anzutreffen, die mindestens zum Teil nicht in eine evolutionäre Abfolge gebracht werden können.

Abb. 4: Ungewöhnliche Federtypen bei (v. l. n. r.): Anchiornis, Epidexipteryx, Beipiaosaurus und Cruralispennia. (Quellen v. l.: Nach Saitta et al. 2017; LWL-Museum für Naturkunde; Matt Martyniuk, CC BY-SA 3.0; Wang et al. 2017; CC SA 4.0) 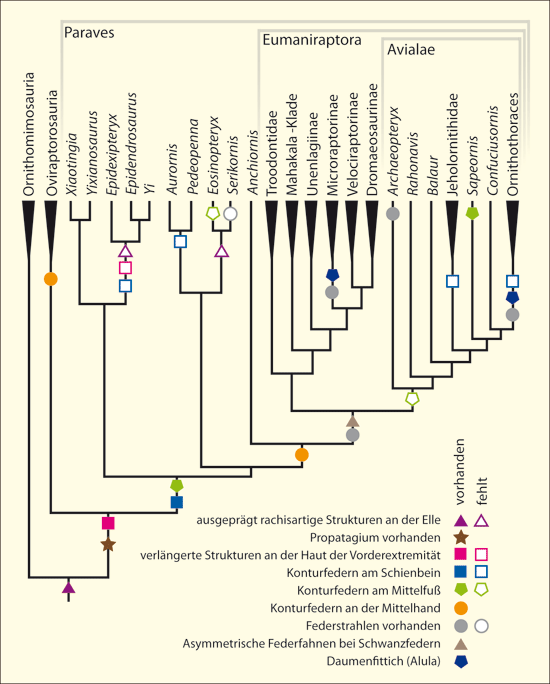
Abb. 5: Cladogramm mit Angabe von Federtypen auf der Basis der phylogenetischen Analyse von Lefèvre et al. (2017). Der Baum offenbart zahlreiche Homoplasien (Konvergenzen oder Rückentwicklungen).
Legt man die vielen Bauplanunterschiede unter den geflügelten Paraves* und den in Oberjura und Unterkreide überlieferten Vögeln zugrunde, folgt daraus gemäß evolutionstheoretischen Prinzipien, dass der aktive Flug nicht nur einmal, sondern in verschiedenen Gruppen unabhängig entstanden ist (Brusatte 2017b, 793; Wang et al. 2019)10 – vor nicht allzu langer Zeit wäre die Möglichkeit einer mehrfach unabhängigen Entstehung des aktiven Vogelflugs evolutionstheoretisch noch ausgeschlossen worden.
Dazu kommt, dass gerade am Anfang der mutmaßlichen Evolution der Vögel eine große Vielfalt von Federtypen fossil überliefert ist, auch solche, die sonst und unter heutigen Vögeln nicht bekannt sind. Das gilt z. B. für den Troodontiden Anchiornis, der u. a. einen Federtyp besaß, der weder typisch daunenartig war noch in seiner Form typische Kennzeichen einer Konturfeder aufwies und auch nicht intermediär zwischen diesen beiden Federtypen ausgeprägt war (Saitta et al. 2017, Abb. 4a). Zudem war das gesamte Federkleid von Anchiornis ebenso einzigartig wie dasjenige der jüngst entdeckten Gattung Serikornis (Lefèvre et al. 2017) aus derselben Familie mit wiederum anderem Merkmalsmosaik bezüglich der Befiederung, das evolutionstheoretisch schwer eingeordnet werden kann.11
Weiterhin wurden bei mehreren Gattungen des Oberjura und der Unterkreide bandartige Federn entdeckt (Abb. 4b), die so unsystematisch verteilt sind, dass evolutionstheoretisch eine mindestens vier Mal unabhängige Entstehung dieses Federtyps angenommen werden muss (Xu et al. 2010).Bei Protopteryx waren die Schwanzfedern ungewöhnlich und einmalig ausgebildet: Sie waren im körpernahen Bereich unverzweigt (Zhang & Zhou 2000, 195712). Weitere Federtypen sind verlängerte breite fadenförmige Federn beim Therizinosaurier Beipiaosaurus (Xu et al. 2009, Abb. 4c) und eine bisher unbekannte Ausprägung bei der Unterkreide-Gattung Cruralispennia aus der Gruppe der Gegenvögel*. Deren Federn waren im körpernahen Bereich drahtartig und besaßen distal fädige Spitzen (Wang et al. 2017; Abb. 4d). Eine größere Vielfalt von Federtypen als heute war früh etabliert13 und trat recht abrupt auf. Und sie lässt sich nicht ohne Weiteres in ein evolutionstheoretisches Schema einfügen. Auch andere Gefiedermerkmale treten teils unsystematisch auf und erfordern die Annahme von Konvergenzen (Beingefieder, gegabelte Schwanzfedern; Daumenfittich).
Eine größere Vielfalt von Federtypen als heute war früh etabliert und trat recht abrupt auf.
Aktuelle Cladogramme* (Abb. 5) verdeutlichen die unsystematische Verteilung verschiedener Federtypen und Arten der Befiederung. Eine enorme Vielfalt von Schwanztypen kam bei Theropoden-Gattungen und Vögeln der Unterkreide relativ zeitgleich vor und kann nur mit vielen Konvergenzen und / oder Rückentwicklungen in evolutionäre Abfolgen eingeordnet werden.
Zähne und Schnabel
Ein herausragendes Merkmal der Vögel ist der Schnabel. „Abgesehen von den Federn ist der Schnabel sicherlich das typischste vogelähnliche Merkmal des Vogelkörpers“14 (Proctor & Lynch 1993, 66). Vögel benutzen ihre Schnäbel nicht nur zur Nahrungsaufnahme, sondern auch zur Pflege ihres Gefieders, zum Nestbau, zur Verteidigung und bei der Balz. Die knöcherne Grundlage des Schnabels – Ober- und Unterkiefer – ist mit einer Hornsubstanz aus Keratin überzogen, die als Rhamphotheca bezeichnet wird. Der Oberschnabel wird von der Maxilla (Oberkiefer) und anderen Schädelknochen unterstützt und ist über eine Art Scharniergelenk mehr oder weniger beweglich.
Zitate zu Konvergenzen bei Schnäbeln finden sich im Online-Zusatzmaterial unter dino-voegel.pdf
Während alle heutigen Vögel komplett zahnlos sind15, besaßen die meisten fossilen Vögel, die in mesozoischen Schichten (Jura, Kreide) überliefert sind, einen bezahnten Kiefer oder eine Kombination aus Schnabel und Zahnkiefer (z. B. Hesperornis, Martyniuk 2012, 43). Die Rhamphotheca weist auch bei fossilen Formen in keinem bekannten Fall Alveolen (Vertiefungen für Zähne) auf; bei Zahnvögeln, die neben den Zähnen auch einen Schnabel besitzen, sind Schnabel und Zähne klar auf verschiedene Teile des Kiefers verteilt und dienen verschiedenen Aufgaben bei der Nahrungsaufnahme; der manchmal verwendete Ausdruck „Zähne im Schnabel“ ist falsch (Martyniuk 2012, 43).16
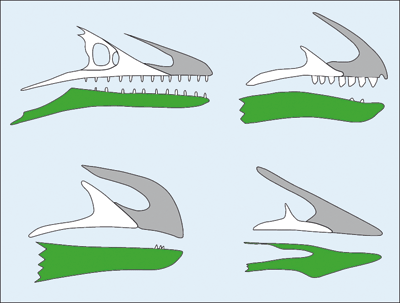
Abb. 6: Vergleich der Bezahnung bei (von oben links) Archaeopteryx lithographica, Sapeornis chaoyangensis, Jeholornis prima und Confuciusornis sanctus. Prämaxilla grau, Maxilla weiß, Dentale grün. (Nach Wang et al. 2017b)
Die Ausprägung der Zähne bei bezahnten Vögeln ist variabel, offenbar in Abhängigkeit von der Art der Ernährung (Louchart & Viriot 2011, 663). Die Vielfalt der Ausprägungen der Zähne bei Zahnvögeln zeigt, dass es sich nicht um bloße Rückbildungsstadien auf einem Weg zum zahnlosen Hornschnabel handelt, sondern um unterschiedlich spezialisierte Ausprägungen.
Zähne und Schnäbel bei Kreidevögeln. Die in der Kreide verbreiteten Gegenvögel* waren meist voll bezahnt ohne Ansätze eines Schnabels, einzige Ausnahme ist die völlig zahnlose Gattung Gobipteryx aus der Oberkreide der Süd-Mongolei, die einen Schnabel besaß (Martin & Zhou 1997). Dagegen besaßen die Ornithuren* kleine Schnäbel am Vorderende des Kiefers und Zähne im hinteren Teil (Martyniuk 2012, 43f.).
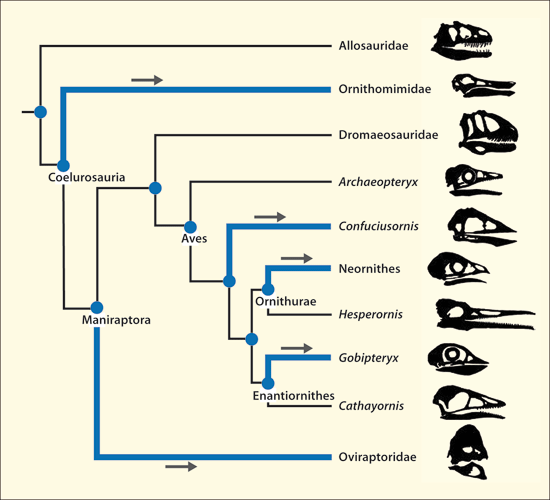
Abb. 7: Cladogramm der Hauptlinien der Coelurosaurier-Theropoden, das fünf unabhängige Fälle von Zahnverlust zeigt. Bei Vögeln gingen die Zähne in mindestens drei Linien unabhängig voneinander verloren: bei den Confuciusornithiden, beim Enantiornithinen Gobipteryx minuta und bei den Neornithes. (Nach Chiappe et al. 1999, 70) Wang et al. (2017) nehmen sogar siebenmalige unabhängige Entstehung von Zahnverlust an.
Bemerkenswerterweise hat ausgerechnet Archaeorhynchus, der an der Basis der Ornithuren steht (Wang & Zhou 2016; Zhou & Zhang 2006, 367), einen völlig zahnlosen Kiefer und einen flachen, löffelförmigen Schnabel.17 Das trifft ebenfalls auf den als noch „primitiver“ eingeschätzten Confuciusornis zu (Abb. 6). Auch die komplett zahnlose Ornithuren-Gattung Hongshanornis steht relativ basal und gehört zu den ältesten Vertretern dieser Gruppe (Zhou & Zhang 2005). Schließlich sei der an der Basis der Vögel stehende Jeholornis erwähnt, der nur drei kleine Zähne am Unterkiefer besaß (Zhou & Zhang 2003a; Abb. 6).
Zahnreduktion und Schnäbel bei Theropoden. Eine Zahnreduktion bis hin zu völliger Zahnlosigkeit findet sich nicht nur bei den kreidezeitlichen Vogelgruppen, sondern auch bei einer Reihe von Gruppen von „Nicht-Vogel-Theropoden“. Evolutionstheoretisch wird eine unabhängige Zahnreduktion bei den Oviraptorosauria, Ornithomimosauria und Therizinosauria angenommen. Während bei basalen (und stratigraphisch älteren) Vertretern Zähne vorkommen, sind viele stratigraphisch jüngere Formen zahnlos oder nur teilweise bezahnt (Zanno 2010; Lautenschlager et al. 2013, 20657).18
Zahnlos war weiterhin auch Zhongornis haoae, eine juvenile Art unklarer systematischer Zugehörigkeit, die in den Bereich der Basis der Vögel gestellt wird (O’Connor & Sullivan 2014).
Schließlich sei noch Limusaurus erwähnt. Diese Gattung der Ceratosauria besaß einen voll entwickelten Schnabel und war komplett zahnlos, wird aber zu den basalen Theropoden und – anders als die anderen zahnlosen Formen – nicht zu den Coelurosauriern und damit nicht in die nähere Verwandtschaft zu den Vögeln gestellt (Xu et al. 2013). Wie bei den anderen Gruppen muss eine unabhängige Entstehung des Schnabels angenommen werden (Abb. 7).
Schlussfolgerungen. Zahnlose Formen oder Gattungen mit einer reduzierten Anzahl von Zähnen sind so unsystematisch im System der Theropoden und kreidezeitlichen Vögel verteilt, dass eine vielfach unabhängige Entstehung von Schnäbeln mit Rhamphotheca bzw. Zahnreduktion angenommen wird (Abb. 7; vgl. Zitate Online-Zusatz). Während innerhalb einzelner Gruppen (Ornithomimosauria, Oviraptorosauria, Therizinosauria, Enantiornithes) grobe Trends Richtung Zahnlosigkeit erkennbar sind, stören Confuciusornis, Archaeorhynchus und Zhongornis dieses Bild erheblich. Denn diese Gattungen gehören zu den stratigraphisch ältesten Formen mit einem Schnabel und dieses Merkmal tritt unvermittelt auf. Archaeorhynchus ist zudem nach phylogenetischen* Analysen ausgerechnet an der Basis der Ornithuromorpha statt wie aufgrund der Ausbildung eines Schnabels eher zu erwarten an einer abgeleiteten* Position.
Die Verteilung der zahnlosen Formen im System widerspricht in Bezug auf das Merkmal „Schnabel“ der These, dass Vogelmerkmale bereits in den Dinosaurier-Vorläufern entstanden seien. Denn die Schnäbel bei Dinosauriern können evolutionstheoretisch nur als konvergente Bildungen interpretiert werden, die unabhängig entstanden und zudem stratigraphisch meist deutlich später als die ältesten komplett zahnlosen Vogelgattungen fossil überliefert sind (Archaeorhynchus, Confuciusornis, Hangshanornis). Mit Confuciusornis tauchen Vogelschnäbel plötzlich in voll ausgeprägter Form auf und sie existierten zeitgleich mit anderen Formen, die einen Zahnkiefer besaßen.
Furkula
Ein wichtiges vogeltypisches Merkmal ist die Furkula (Gabelbein). Sie wird mit den beiden Schlüsselbeinen homologisiert, die miteinander zu einem V- oder U-förmigen bzw. beidseitig sigmoiden Gebilde verwachsen sind (Abb. 8). Bei heutigen Vögeln variiert die Furkula erheblich in Größe, Form, Steifheit und Orientierung zum Rest des Schultergürtels (Close & Rayfield 2012, 119); ihre Funktion kann entsprechend ebenfalls variieren. Die Unterschiede hängen tendenziell, aber nicht ausschließlich mit der Art des Flugs zusammen.

Abb. 8: Furkulas bzw. Schlüsselbeine einiger Theropoden und der Taube (Columba). Bei Coleophysis liegen getrennte Schlüsselbeine vor. (Nach Padian & Chiappe 1998)
Die Funktion der Furkula als elastische, energiespeichernde Spange erfordert viele Abstimmungen im Vogelkörper.
Der Furkula wird die Funktion einer gespannten Feder bzw. elastischen Spange zwischen den Schultergelenken zugeschrieben, die Energie beim Flügelschlag speichert. Die Furkula dient außerdem als Verstärkung des Brustskeletts zur Unterstützung bei den Belastungen während des Flugs und als Ansatz für die Flugmuskeln, vor allem beim Abschlagen der Flügel. Sie stabilisiert auch das Schultergelenk und ist mit den beiden Schulterblättern verbunden. Außerdem wird der Furkula eine Funktion bei Atmungsbewegungen zugeschrieben (Jenkins et al. 1988; Nesbitt et al. 2009, 859; Bock 2013, 123620).
Bei fossilen Vögeln war die Funktionalität der Furkula wahrscheinlich teilweise anders als bei den heutigen Vögeln. So vermuten Olson & Feduccia (1979), dass bei Archaeopteryx die Furkula gleichsam das schwach ausgebildete Brustbein als Ansatzstelle für die Flugmuskulatur teilweise ersetzte, was Bock (2013, 1238) allerdings in Frage stellt.
Homologie. Die Homologie von Furkula und Schlüsselbeinen wird begründet mit der ähnlichen Lage, weiter damit, dass Furkula und Schlüsselbeine nie gemeinsam auftreten und dass beide eine ähnliche Skelettentwicklung durchlaufen (Hall & Vickaryous 2015, 440).21 Diese Homologie ist jedoch nicht unumstritten (Bryant & Russell 1993; Feduccia 1999, 77; Hall & Vickaryous 2015). Feduccia hält die Architektur der Schulter von Dinosauriern für so dramatisch verschieden von der von Archaeopteryx und von modernen Vögeln, dass es unwahrscheinlich sei, dass einer der Schulterknochen in ähnlicher Weise mit der Furkula verbunden gewesen sei und eine ähnliche Funktion ausgeübt habe wie bei Vögeln.22 Auf der Basis eines Überblicks über Theropoden mit nachgewiesener Furkula haben Nesbitt et al. (2009, 875) jedoch keinen Zweifel mehr daran, dass die Furkula der Vögel mit den Schlüsselbeinen anderer Tetrapoden (Vierbeiner) homolog sei; das folge sowohl aus phylogenetischen als auch entwicklungsbiologischen Daten.23 Hall & Vickaryous (2015) bestreiten später wiederum aufgrund ihrer eigenen Studien, dass die Homologie gesichert sei. Die Furkula könnte auch mit der Interclavicula (Verbindung Schlüsselbein-Brustbein) homolog sein. Diese beiden Autoren kommen zum Schluss (S. 150): „Wir wissen immer noch nicht, ob die Furkula der Interclavicula entspricht, eine Neubildung ist oder verschmolzene Schlüsselbeine darstellt.“24
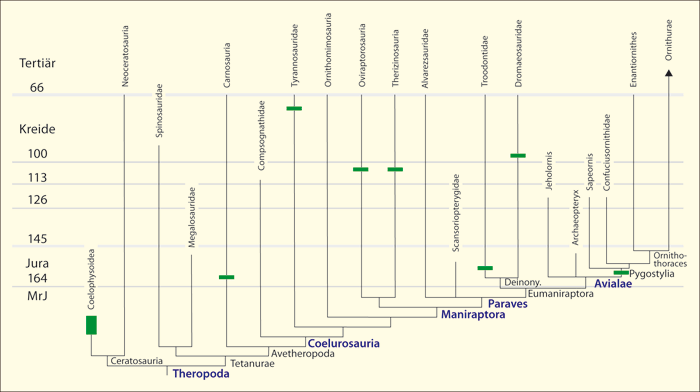
Abb. 9: Verteilungen von Theropoden- und Vogelgattungen mit Furkula. (Nach mehreren Quellen zusammengestellt; s. Junker 2019)
Verteilung. Nachdem der Besitz einer Furkula lange Zeit ein exklusiv vogelspezifisches Merkmal zu sein schien, hat sich jedoch nach und nach herausgestellt, dass die Ausbildung einer Furkula bei Theropoden-Dinosauriern verbreitet war (vgl. Abb. 3). Daher wird heute angenommen, dass die Furkula bei den Vögeln für den Flug kooptiert wurde, nachdem sie zuvor eine andere Funktion innehatte (Brusatte 2017a, 54).
Nesbitt et al. (2009) geben einen Überblick über die Theropodengruppen, bei denen eine Furkula nachgewiesen wurde und kommen zu dem Schluss, dass alle größeren Linien der Theropoden von Beginn an eine Furkula besaßen (plesiomorphes* Merkmal); der Besitz einer Furkula sei daher kennzeichnend für die Theropoden insgesamt.25 Das Fehlen einer Furkula bei den Ornithomimosauria und Alvarezsauridae könnte auf mangelhafte fossile Erhaltung zurückgeführt werden26 und wäre sonst am ehesten als sekundärer Verlust interpretierbar.27 Alternativ müsste eine vielfach konvergente Entstehung einer Furkula angenommen werden, was Tykoski et al. (2002, 730f.) nicht ausschließen.28
Bemerkenswert ist, dass auch bei den Scansoriopterygiden, die in die Nähe der Vögel gestellt werden, keine Furkula, sondern zwei separate Schlüsselbeine nachgewiesen wurden (Czerkas o. J., Czerkas & Yuan 2002, 6). Aufgrund der phylogenetischen Position (s. Abb. 3) müsste bei dieser Gruppe die Furkula in Schlüsselbeine zurückevolviert sein – ein unplausibles Szenario.
Diskussion. In die Reihe der vogeltypischen Merkmale, die Schritt für Schritt bereits bei den Theropoden evolviert sind, lässt sich die Furkula kaum einfügen. Folgt man den phylogenetischen Analysen, war die Furkula schon an der Basis der Theropoden ausgebildet und wäre daher bezüglich genauerer Verwandtschaftsbeziehungen von Theropoden und Vögeln nicht aussagekräftig. Evolutionstheoretisch kann die Ausbildung einer Furkula jedoch als „Voranpassung“ interpretiert werden.
Interessanter ist die Frage, wie ausgehend von einer Theropoden-Furkula die Verhältnisse bei den Vögeln evolvierten. Die Funktion als elastische, energiespeichernde Spange erfordert viele Abstimmungen im Vogelkörper; das gilt erst recht, wenn man bedenkt, dass die Furkula mehrere Funktionen innehat (s. o.). Ein Funktionswechsel bzw. die Integration neuer Funktionen führen ungerichtete evolutionäre Mechanismen an ihre Grenzen. Welche Hürden dabei zu überwinden gewesen wären, müsste ein genauerer Vergleich von Formen zeigen, die in eine evolutionäre Linie gebracht werden. Angesichts der Lückenhaftigkeit der Fossilüberlieferung bleibt hier vieles offen.
Die Homologie der Furkula verschiedener Gruppen ist wie erwähnt nicht gesichert. Dass bei den Scansoriopterygiden, Ornithomimosauria und Alvarezsauriden trotz teilweise guter Erhaltung keine Furkula gefunden wurde, ist evolutionstheoretisch problematisch, da unverständlich ist, dass eine Furkula wieder aufgegeben wird. Bekannt ist dieser Fall bei heutigen sekundär flugunfähigen Vögeln, doch wird man bei den drei genannten fossilen Gruppen kaum einen Flugverlust annehmen wollen. Die Alternative wäre einmal mehr eine mehrfache unabhängige Entstehung einer Furkula.
Die Rückbildung der Furkula bei heutigen flugunfähigen Vögeln wertet Feduccia als Indiz dafür, dass die Furkula eine Neubildung erst bei den Vögeln sei. Er argumentiert: „Wenn dieser [Verlust] ein allgegenwärtiges Merkmal flugunfähiger Vögel ist, warum sollte man dann erwarten, eine voll entwickelte Furkula in flugunfähigen zweibeinigen Dinosauriern zu finden?“ (Feduccia 1999, 26529). Da eine Furkula bei diesen Formen aber vorkommt, müsse sie eine andere Funktion als bei Vögeln gehabt haben, was gegen ihre Homologie mit der Vogel-Furkula spreche.
Insgesamt spricht trotz Feduccias Einwand zwar viel für eine Homologie der Furkula von Vögeln und Theropoden-Dinosauriern, doch kann dies nicht als gesichert gelten. Von der Homologisierbarkeit wiederum hängt es ab, ob die Furkula von Vögeln als Merkmal interpretiert werden kann, das bereits bei den nichtfliegenden mutmaßlichen Vorfahren ausgebildet war.
Pygostyl
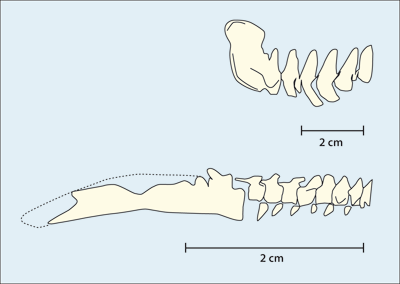
Abb. 10: Pygostyl der Taube (oben) und des Gegenvogels* Iberomesornis. (Nach Gatesy & Dial 1996)
Bei heutigen Vögeln sind die letzten Schwanzwirbel zu einem sogenannten Pygostyl* verwachsen (Abb. 10). Es dient als stabile Unterlage für die fächerförmig angeordneten Schwanzfedern. Das Pygostyl ist beidseitig mit einem „Steuerfederkolben“ verbunden, einem komplizierten Organ aus Fett, Bindegewebe und Muskeln, an dem die Schwanzfedern ansetzen und durch das diese bewegt und gesteuert werden können; nur die zwei mittleren Schwanzfedern sind direkt mit dem Pygostyl verbunden.30 Die damit verbundene Fähigkeit, die Form des Schwanzes zu verändern, verbessert erheblich die Flugfähigkeit (O’Connor et al. 2015, 114; Gatesy & Dial 1996, 2045ff.). Das ganze System bildet eine komplizierte integrierte Einheit und ermöglicht zusammen mit den Flügeln während des Flugs die Ausbildung einer eng gekoppelten Fläche (Gatesy & Dial 1996, 2037f.).31
Wang & O’Connor (2017, 291) sprechen von einem „ausgeklügelten Schwanzkomplex“ und von einer „integrierten Ganzheit“; das Uropygium (Steißgegend) und Integument (Körperbedeckung) stehen in einer morphologischen Wechselbeziehung, so dass von der Form des einen die des anderen vorhergesagt werden könne. Diese enge Wechselbeziehung stütze die evolutionstheoretische Hypothese, dass Pygostyl und Schwanzfedern co-evolvieren.32
Auch bei den meisten fossilen Vögeln ist ein Pygostyl ausgebildet; sie werden als Pygostylia* zusammengefasst, wobei es bei basalen Arten deutlichere Unterschiede in der Ausprägung im Vergleich zu heutigen Arten gibt. Dagegen ist das Skelett der Schwanzregion heutiger Vögel trotz Vielfalt der Schwanzausprägungen bemerkenswert einheitlich.
Pygostyltypen bei fossilen Formen. Unter den frühen Vögeln, die im geologischen System der Kreide fossil überliefert sind, sind drei verschiedene Pygostyltypen überliefert, von denen unter den heutigen Vögeln nur der erste vorkommt (Hu et al. 2015, 16f.; Wang & O’Connor 2017, 30433): 1. kurz, pflugförmig und sich verjüngend34, 2. kräftig, gabelig und körperfern eingeschnürt und 3. kräftig und stabartig.
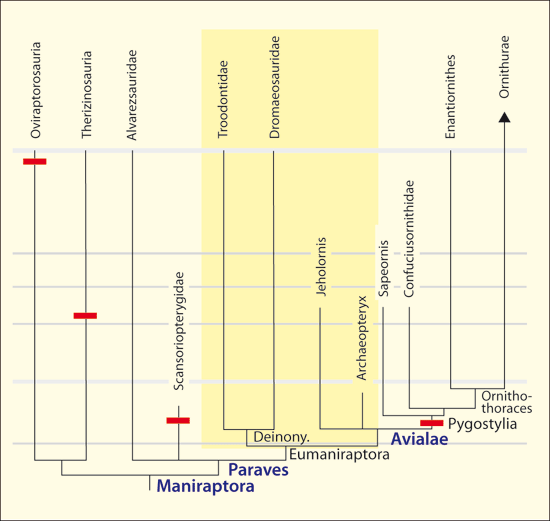
Abb. 11: Verteilungen von Theropoden- und Vogelgattungen mit Pygostyl. Man beachte, dass bei den Gattungen, die an der Basis der Vögel stehen, kein Pygostyl nachgewiesen wurde. Die Theropoden-Dinosaurier, die ein Pygostyl besitzen, sind fossil viel später überliefert als die ältesten Vögel mit Pygostyl (Pygostylia). (Nach zahlreichen Quellen zusammengestellt; s. Junker 2019)
Die zweibeinigen Theropoden besaßen dagegen in der Regel lange Schwänze mit Dutzenden von freien Schwanzwirbeln, die mutmaßlich der Balance dienten. Robuste quer gerichtete Fortsätze an den vorderen Schwanzwirbeln dienten als Ansatzpunkte für Muskeln, die primär als Rückziehmuskeln der Hinterbeine dienten (Rashid et al. 2014, 2).35 Die beiden Schwanztypen – Fiederschwanz und Fächerschwanz – sind funktionell sehr verschieden. Die Hinterbeine waren funktionell mit dem langen Schwanz verbunden (Benson & Choiniere 2013, 1). Zur Entwicklung des Fächerschwanzes musste diese funktionelle Kopplung aufgehoben werden, was zusätzlich unterstreicht, dass der Übergang zu einem Fächerschwanz einen erheblichen Umbau erforderte. Einige Gattungen der Theropoden-Dinosaurier besaßen aber ebenfalls ein Pygostyl (s. u.).
Verteilung. Vögel mit „Dino-Schwanz“ (Jeholornis und verwandte Arten, Microraptor und Archaeopteryx) und Arten mit Pygostyl und Fächerschwanz lebten zeitweise nebeneinander (Rashid et al. 2018, 8). Viele Maniraptoren (mit langen Schwänzen), die als Vorläufergruppen von Vögeln gelten, sind stratigraphisch (z. T. deutlich) jünger als zahlreiche Pygostylia. Die Arten mit Pygostyl oder pygostylartigen Strukturen sind im Theropoden-Cladogramm in einer Weise auf verschiedenen Ästen verteilt, dass auch hier eine mehrfach unabhängige (konvergente) Entstehung angenommen werden muss (Abb. 10).
Wang & O‘Connor (2017, 303) halten es aufgrund der Struktur des Pygostyls bei Oviraptorosauriern für unwahrscheinlich, dass ein Steuerfederkolben oder äquivalente Strukturen ausgebildet waren.36 Persons et al. (2014, 553, 562, 564) sind der Auffassung, dass die Schwanzregion der Oviraptorosauria einzigartig sei und nicht in eine Übergangsposition zu einem vogeltypischen Pygostyl passe, sondern eine konvergente Bildung sei und am ehesten eine Schaufunktion gehabt habe.37 Dass die unterschiedlichen Pygostyle als konvergent interpretiert werden müssen, ergibt sich auch aus ihrer Verteilung im Cladogramm (s. o.), es sei denn man interpretiert die Oviraptorosauria als sekundär flugunfähige Vögel.
Übergangsformen? Wang & O’Connor (2017, 289) bezeichnen die Entstehung des Pygostyls und der fächerförmigen Anordnung der Federn als eine der „bemerkenswertesten Anpassungen“ der frühen Vogelevolution, doch ein allmählicher Übergang zwischen Formen ohne und mit einem Pygostyl sei fossil nicht dokumentiert (auch O‘Connor et al. 2015, 11437 Mayr 2017, 4538, Wang & Zhou 2017, 639). Die ältesten Pygostyle aller Ornithuromorpha aus der Unteren Kreide sind nahezu „modern“ ausgebildet. Zusammen mit der Überlieferung fächerförmiger Schwanzfedern spricht dies evolutionstheoretisch dafür, dass der komplette Vogelschwanz-Komplex sehr früh in dieser Linie evolvierte und dass die Steuerfederkolben und der Fächerschwanz gemeinsam evolvierten (so nach Wang & O’Connor 2017, 289, 30540 sowie O’Connor et al. 2015, 114). Es sei die sparsamste Erklärung, dass der Steuerfederkolben bei den Confuciusornis-Verwandten und einigen Gegenvögeln verloren gegangen sei (O’Connor et al. 2015, 11441). Dagegen argumentieren O’Connor et al. (2015, 117), dass es unverständlich sei, dass die abgeleiteten Gruppen der Gegenvögel diesen Komplex zugunsten eines robusteren Pygostyls wieder aufgegeben hätten und plädieren für eine unabhängige Entstehung der Pygostyle mit dem Steuerfederkolben; das werde auch dadurch unterstützt, dass es morphologische Unterschiede bei den Pygostylen gibt.42
Als Zwischenform wurde zwar Zhongornis haoae (eine juvenile Art unklarer systematischer Zugehörigkeit) diskutiert, doch zeigte eine erneute Untersuchung, dass ungefähr 20 Schwanzwirbel erhalten waren (O’Connor & Sullivan 2014). Es ist auch möglich, dass das Pygostyl noch nicht ausgereift war (Rashid et al. 2018).43
Fazit. Das Pygostyl gehört nicht zu den vogeltypischen Strukturen, die bereits bei dinosaurierartigen Vogelvorläufern evolviert sind. Zwar gibt es eine konvergente Entstehung bei jüngeren Formen von Theropoden-Dinosauriern, die aber erst deutlich nach dem Auftreten der ersten Vögel fossil überliefert sind (Abb. 10), folglich können sie nicht als Vorläufer von Vögeln interpretiert werden. Es sind zwar verschiedene Pygostyltypen bekannt, auch solche, die unter heutigen Vögeln nicht vorkommen, aber Formen mit und solche ohne Pygostyl (und folglich mit Fächerschwanz bzw. Fiederschwanz) sind klar voneinander getrennt. Es gibt jedoch einige Vogelgattungen mit einem Fiederschwanz, nämlich Jeholornis und Archaeopteryx sowie – falls man diese Gattung zu den Vögeln rechnet – der vierflügelige Microraptor.
Deutung aus der Schöpfungsperspektive
Geht man statt von Makroevolution von einer Erschaffung von Grundtypen? aus, kann man die diskutierten Befunde wie folgt interpretieren:
• Das explosive Auftreten der Formenvielfalt spiegelt die Existenz von Grundtypen wieder.
• Die zahlreichen, unterschiedlichsten Mosaikformen sind Ausdruck vielfältiger Kombinationen von Merkmalen, deren Ausprägung durch die jeweilige Lebensweise und nicht durch eine vorausgegangene Evolution erklärt wird. Ein Schöpfer ist frei in seinem Handeln, während Evolution an einen Mechanismus gebunden ist.
• Die Schwierigkeiten, die sich aus der Existenz verschiedenster Mosaikformen (häufige Konvergenzen) ergeben, erübrigen sich, wenn die Merkmale frei und entsprechend der Anforderungen für bestimmte Lebensweisen und Lebensräume kombiniert werden können.
• Die Problematik einer „Experimentierung“ entfällt. Es gibt keine „Experimente“, sondern eine anfängliche Vielfalt von Formen, die ursprünglich in mancher Hinsicht am größten war (besonders bei Federtypen und Flugformen).
• Die Frage nach Evolutionsmechanismen für schnelles und vielfältiges Auftreten verschiedenster Formen entfällt.

Tab. 1: Überblick zur Analyse der Verteilung vogeltypischer Merkmale bei Theropoden-Dinosauriern. Es fällt auf, dass viele der untersuchten Vogelmerkmale nicht bei denjenigen Formen ausgebildet sind, die als unmittelbare Vogelvorläufer angesehen werden, sondern konvergent entstanden sein müssen (gelbe Unterlegung). Ein Großteil der anderen untersuchten Merkmale gilt als allgemeines Theropodenmerkmal (hellrote Unterlegung) und ist daher ebenfalls nicht gut geeignet, eine schrittweise Entstehung von Vogelmerkmalen bei Theropoden-Dinosauriern zu belegen. Allgemeine Schlussfolgerungen aus den Einzelstudien
Einige Befunde sind evolutionstheoretisch gut interpretierbar. Manche Mosaikformen könnten evolutionären Übergangsformen nahestehen. Die meisten Mosaikformen eignen sich dafür aber nicht, weil ihre Merkmalskombinationen insgesamt nicht in basale Positionen von Formengruppen passen. In einigen Gruppen können Tendenzen nachvollzogen werden (z. B. verschiedene Ausprägungen von Schnäbeln und verschiedenes Ausmaß an Zahnreduktionen; unterschiedliche Pygostyltypen).44 Auch die Tatsache, dass viele Vogelmerkmale bei verschiedenen Dinosaurier-Gruppen der Theropoden vorkommen, kann als Punkt für evolutionäre Deutungen gewertet werden, allerdings mit den diskutierten Einschränkungen.

Tab. 2: Gegenüberstellung evolutionstheoretischer Vorhersagen bzw. Erwartungen und Beobachtungen am Fossilbefund.
Die Einzeluntersuchungen (auch der hier nicht angeführten Merkmale [Junker 2019]) haben jedoch gezeigt, dass die vogeltypischen Merkmale so im System der Theropoden und Kreidevögel verteilt sind, dass fast durchweg Konvergenzen oder Reversionen angenommen werden, meist mehrfach. Evolutionstheoretisch bedeutet das, dass diese Merkmale zwei- bis mehrfach unabhängig entstanden sein müssen. O’Connor et al. (2011, 40f.) kommen zum Schluss, dass die Verteilung vogelartiger Merkmale wie Schnabel, Furkula, Brustbeinplatten, hakenförmige Rippenfortsätze, rückwärtsgerichtetes Schambein, distal nicht verwachsene Schambeine, Parallelität von Schambein und Sitzbein und die Ausbildung eines Pygostyls in einer Weise innerhalb der Maniraptoren verbreitet seien, „dass es für die meisten von ihnen schwierig ist, ein klares Muster der Entstehung der Merkmale zu bestimmen.“ Zahlreiche Autoren merken an, dass es unübersichtliche Verwandtschaftsverhältnisse gebe und Konvergenzen verbreitet seien (vgl. Tab. 1 und Zitate im Online-Zusatzmaterial). In einigen Fällen müsste man unter evolutionstheoretischen Vorgaben annehmen, dass „Vogelmerkmale bei Dinosauriern“ nach ihrer Entstehung wieder verloren gegangen sind, bevor Vögel evolviert sind. In solchen Fällen können sie folglich keine Vorläufermerkmale sein.
Die Behauptung, Vögel stammten von Dinosauriern ab oder seien sogar Dinosaurier, ist durch die fossilen Daten nicht gedeckt.
Diese Situation ist umso bemerkenswerter, als aufgrund des Fehlens jeglicher Zielorientierung in der Evolution Konvergenzen nicht zu erwarten sind und erst recht nicht ein gehäuftes Auftreten. Insgesamt zeigt sich, dass ein schrittweiser Erwerb von Vogelmerkmalen in Dinosauriergruppen anhand des fossilen Befundes nur in sehr eingeschränktem Sinne nachvollziehbar ist. Nur eine summarische Darstellung ohne Beachtung der Details kann diesen Eindruck erwecken. Die Behauptung, Vögel stammten von Dinosauriern ab oder seien sogar Dinosaurier, ist durch die fossilen Daten nicht gedeckt.
Weitere Schlussfolgerungen sind in Tab. 2 zusammengefasst, in der die Erwartungen im Rahmen einer Makroevolution den fossilen Befunden gegenübergestellt werden.
Die komplette Literaturliste findet sich im Online-Zusatzmaterial unter dino-voegel.pdf.
Bock WJ (2013)
The furcula and the evolution of avian flight. Paleont. J. 47, 1236–1244.
Brusatte SL, Lloyd GT, Wang SC & Norell MA (2014)
Gradual assembly of avian body plan culminated in rapid rates of evolution across the dinosaur-bird transition. Curr. Biol. 24, 2386–2392.
Brusatte SL (2017a)
Taking wing. Sci. Am. Jan. 2017, 48–55.
Brusatte S (2017b)
A Mesozoic aviary. Science 355, 792–794.
Feduccia A (1999)
The origin and evolution of birds. Yale University Press. 2nd ed.
Junker R (2017)
Dino-Federvieh. Zum Ursprung von Vogelfeder und Vogelflug. W+W Special Paper. https://www.wort-und-wissen.org/wp-content/uploads/b-17-1_feder-und-flug.pdf.
Junker R (2019)
Sind Vögel Dinosaurier? Eine kritische Analyse fossiler Befunde. W+W Special Paper. https://www.wort-und-wissen.org/wp-content/uploads/b-19-4_dinos-voegel.pdf.
Lautenschlager S, Witmer LM, Altangerel P & Rayfield EJ (2013)
Edentulism, beaks, and biomechanical innovations in the evolution of theropod dinosaurs. Proc. Natl. Acad. Sci.110, 20657–20662.
Louchart A & Viriot L (2011)
From snout to beak: the loss of teeth in birds. Trends Ecol. Evol. 26, 663–673.
Martyniuk MP (2012)
A field guide to Mesozoic birds and other winged dinosaurs. Vernon, NJ: Pan Aves.
Mayr G (2017)
Avian evolution. Wiley, Chichester.
O’Connor J, Chiappe LM & Bell A (2011)
Pre-modern birds: Avian divergences in the Mesozoic. In: Dyke G & Kaiser G (eds) Living Dinosaurs: The Evolutionary History of Modern Birds. John Wiley & Sons, pp 39–114.
Padian K & Chiappe LM (1998)
The origin of birds and their flight. Sci. Am. 2/1998, 38–47.
Proctor NS & Lynch PJ (1993)
Manual of ornithology. Avian structure & function. Yale Univ. Press.
Rashid DJ, Chapman SC, Larsson HCE, Organ CL, Bebin AG, Merzdorf CS, Bradley & & Horner JR (2014)
From dinosaurs to birds: a tail of evolution. Evo-Devo 5:25, http://www.evodevojournal.com/content/5/1/25.
Wang M, O’Connor JK, Pan Y & Zhou Z (2017)
A bizarre Early Cretaceous enantiornithine bird with unique crural feathers and an ornithuromorph plough-shaped pygostyle. Nat. Comm. 8:14141.
Wang W & O’Connor JK (2017)
Morphological coevolution of the pygostyle and tail feathers in Early Cretaceous birds. Vertebr. Palasiatica 55, 289–314.
Witmer LM (2002)
The debate on avian ancestry: Phylogeny, function, and fossils. In: Chiappe LM & Witmer LM (eds) Mesozoic birds: Above the heads of dinosaurs. Berkeley, Los Angeles, London: University California Press, pp 3–30.
Xu X (2006)
Feathered dinosaurs from China and the evolution of major avian characters. Integr. Zool. 1, 4–11.
Xu X, Zhou Z, Dudley R, Mackem S, Chuong CM, Erickson GM & Varricchio DJ (2014)
An integrative approach to understanding bird origins. Science 346, 1341, 1253293-1–1253293-10.
Zhou Z & & Zhang F (2003a)
Jeholornis compared to Archaeopteryx, with a new understanding of the earliest avian evolution. Naturwissenschaften 90, 220–225.
Themen | Kurzbeiträge | Streiflichter
Studiengemeinschaft WORT und WISSEN e.V.
Letzte Änderung: 6/25/21
Webmaster