
Themen • Kurzbeiträge • Streiflichter
Stammt der Mensch von einem bayerischen Menschenaffen ab?
Funktionsmorphologische und evolutionstheoretische Spekulationen
von Michael Brandt
Studium Integrale Journal
27. Jahrgang / Heft 1 - Mai 2020
Seite 4 - 11
Zusammenfassung: Der Ursprung des menschlichen zweibeinigen Ganges ist eine zentrale Frage im Evolutionsparadigma. Diese Fortbewegungsweise soll sich nach der Abspaltung des Schimpansen vom gemeinsamen Vorfahren mit dem Menschen entwickelt haben. Neu entdeckte fossile Überreste eines bisher unbekannten miozänen Primaten aus dem bayerischen Allgäu werden von Forschern nun jedoch als Beleg dafür angeführt, dass diese Evolution bereits bei sich in Bäumen fortbewegenden Großaffen begann. Ist diese Hypothese stichhaltig und wie könnte der neu entdeckte Menschenaffe Danuvius guggenmosi in einem Schöpfungsmodell gedeutet werden?
• • • • • • •
Einleitung
Bei der Erforschung des Ursprungs des Menschen stellt sich im Evolutionsmodell eine entscheidende Frage: Wann und wie hat sich der aufrechte Gang entwickelt? Die gewohnheitsmäßige Fortbewegung auf zwei Beinen wird als das Schlüsselmerkmal von Homininen angesehen. Homininen sind Wesen, die evolutionstheoretisch dem Menschen näherstehen als dem Schimpansen, der als nächster Verwandter des Menschen angesehen wird. Anders gesagt: Im Evolutionsmodell werden alle Wesen auf der phylogenetischen Linie zum Menschen nach der Abspaltung des Schimpansen vom letzten hypothetischen gemeinsamen Vorfahren zusammen mit dem Menschen als Homininen bezeichnet.
Heute wird angenommen, dass der Beginn der gewohnheitsmäßigen zweibeinigen Fortbewegung und damit Evolution des frühesten Homininen am Ende des Miozäns vor 5-7 Millionen Isotopenjahren stattfand (Kivell 2019).
Welche Art der Fortbewegung hatte der letzte gemeinsame Ahne von Mensch und Schimpanse? Evolvierte die regelmäßige zweibeinige Fortbewegung auf dem Erdboden ausgehend von einem Ahnen, der hauptsächlich in Bäumen lebte und dann herabstieg (top-down), oder ging dieser hypothetische Ahne auf allen Vieren auf dem Erdboden, stand auf und lief dann auf zwei Beinen (bottom-up)?
Böhme et al. (2019) meinen der Beantwortung dieser Frage entscheidend näher gekommen zu sein. Die Autoren berichten in Nature über den Fund eines bisher unbekannten fossilen Menschenaffen aus dem bayerischen Allgäu, den sie Danuvius guggenmosi nennen (Abb. 1, 2). Die Fossilien wurden aus Ablagerungen mit einem datierten Alter von 11,62 Millionen Jahren geborgen.
Die Veröffentlichung von Böhme et al. (2019) hat große mediale Aufmerksamkeit erfahren. Ob diese auch gerechtfertigt ist, möge der Leser nach dem Studium dieses Artikels selbst beurteilen.
Mit einem Stern* versehene Begriffe werden im Glossar erklärt.
Abb. 1: Die 21 Knochen des Teilskeletts des am vollständigsten erhaltenen (männlichen) Danuvius guggenmosi. https://www.sueddeutsche.de/wissen/danuvius-guggenmosi-allgaeu-menschenaffe-1.4673554
Kompakt
Madelaine Böhme und ihre Kollegen haben Ende 2019 mit Danuvius guggenmosi eine neue fossile miozäne Menschenaffenart beschrieben, die nach den Merkmalen der Zähne und der Knochenüberreste des Schädels aus dem Formenkreis der Dryopithecinen stammt. Der Affe aus dem bayerischen Allgäu wird auf 11,62 Millionen radiometrische Jahre datiert. Die Körperproportionen von Danuvius sind ähnlich denen des Zwergschimpansen. Im Bereich des Körperstamm- und Extremitätenskeletts zeigt der neue Primat aber eine bisher unbekannte Kombination von Merkmalen, die auf eine unbekannte Fortbewegungsweise schließen lässt. Böhme et al. (2019) vermuten, dass es sich dabei um eine Kombination von Hangeln in Bäumen und einer zweibeinigen Fortbewegung mit gestreckten unteren Gliedmaßen gehandelt hat. Danuvius könnte nach den Autoren ein mögliches Modell der Fortbewegung des letzten gemeinsamen Ahnen von Großaffe und Mensch repräsentieren.
Viele fossile Großaffen aus dem Miozän weisen einen Mix von Skelettanpassungen auf, der unähnlich dem der lebenden Großaffen ist. Es lässt sich kaum näher bestimmen, wie sich diese Großaffen fortbewegten und wie viel Zeit sie dabei in Bäumen oder auf dem Erdboden verbrachten. Diese Einschätzung dürfte angesichts der einmaligen Merkmalskonstellation auch auf Danuvius zutreffen.
Falls Danuvius sich tatsächlich biped* mit gestreckten unteren Extremitäten in Bäumen fortbewegt hat, wie Böhme und ihre Kollegen vermuten, entstand gemäß Evolutionsmodell der zweibeinige Gang schon bei den Menschenaffen lange vor Abspaltung der Homininen* vom gemeinsamen Großaffe-Mensch-Vorfahren, und dies geschah auch nicht in Afrika, wie bisher postuliert, sondern in Europa. Die Bipedie wäre damit nicht mehr das entscheidende Identifikationskriterium von Homininen. Die im Evolutionsmodell als frühe Homininen klassifizierten Primaten mit Kletteranpassungen könnten deshalb auch Großaffen mit einem heute nicht mehr zu beobachtenden Fortbewegungsmix gewesen sein.
Im Grundtypkonzept der Schöpfungslehre repräsentiert Danuvius guggenmosi eine ausgestorbene Art eines Menschenaffen-Grundtyps. Innerhalb von Grundtypen sind aufgrund angelegter (latenter) genetischer Programme zahlreiche Skelettmerkmalskombinationen zu erwarten. Menschenaffen, die sich in Bäumen hangelnd und mit gestreckten Beinen biped auf Baumzweigen abstützend fortbewegten, wie Böhme et al. (2019) vermuten, sind in diesem Modell gut denkbar.
Fortbewegung von Danuvius als Ahnenmodell
Die Fortbewegung von Danuvius repräsentiert nach Böhme et al. (2019) ein Modell für den postulierten gemeinsamen Ahnen von Großaffe und Mensch.
Nach Böhme et al. (2019) bewegte sich Danuvius guggenmosi auf eine zuvor unbekannte Art fort, die sie als „extended limb clambering (ELC)“, also Klettern mit gestreckten Gliedmaßen, bezeichnen. Es ist eine Kombination von Suspension (hangelnder Fortbewegung durch Baumkronen) und bipeder (zweibeiniger) Fortbewegung. Diese Art der Lokomotion ist nach Böhme et al. (2019) ein Modell der Fortbewegung des letzten gemeinsamen Ahnen von Schimpanse und Mensch und ein Vorläufer der obligaten (gewohnheitsmäßigen) zweibeinigen Fortbewegung.
Im Folgenden werden die von Böhme et al. (2019) festgestellten Knochenmerkmale von Danuvius und ihre funktionelle Deutung im Rahmen der Fortbewegung dargestellt und einer kritischen Bewertung unterzogen. Außerdem wird der neue Primat im Rahmen bestehender Ursprungsvorstellungen diskutiert.
Knochenüberreste und Fortbewegung nach Böhme et al. (2019)
Die Fossilien von Danuvius umfassen Überreste von mindestens vier Individuen mit einem Teilskelett, das nach Böhme et al. (2019) für die Beschreibung der Extremitäten und der Wirbelsäule sowie Körperproportionen ausreichend ist.

Abb. 2: Darstellung von Danuvius guggenmosi durch den Künstler Velizar Simeonovski im Auftrag von Madelaine Böhme und ihren Kollegen. https://www.sueddeutsche.de/wissen/menschenaffe-danuvius-allgaeu-tuebingen-1.4671000
Danuvius hatte ein Körpergewicht von ca. 17 bis 31 kg und liegt damit zwischen dem des Siamang (Primatenart aus der Familie der Gibbons) und dem des Zwergschimpansen (Bonobo).
Die craniodentalen* Überreste von Danuvius sind ähnlich denen der Dryopithecinen, einer fossilen Menschenaffengruppe aus dem mittleren bis späten Miozän in Europa. Manche Forscher betrachten die Dryopithecinen als Ahnen der Afrikanischen Großaffen.
Das Körperstamm- und Extremitätenskelett von Danuvius guggenmosi ist jedoch einmalig im Vergleich mit bisher bekannten Dryopithecinenfunden, zum einen aufgrund von Funden zweier beinahe vollständig erhaltener Extremitätenknochen – einer Elle und eines Schienbeins – und zum anderen in der Kombination der Knochenmerkmale. Zusätzlich zu den beiden Extremitätenknochen sind vom Skelett einige Wirbel, ein Teil des Oberschenkels sowie Hand- und Fußknochen erhalten. Im Folgenden werden einige grundlegende Aspekte der Knochenmorphologie und Fortbewegung von Danuvius dargestellt.
Danuvius besitzt zum einen generalisierte Extremitätenproportionen ähnlich denen des Zwergschimpansen (Bonobo), zum anderen aber eine einmalige Kombination der Stellung von Knie, oberem Sprunggelenk, Ellenbogen und Handgelenk sowie stark greifende Extremitäten.
Die Kombination der Merkmale des Körperstamm- und Extremitätenskeletts von Danuvius ist einmalig unter den lebenden und fossilen Primaten.
Das von Böhme et al. (2019) für Danuvius postulierte „extended limb clambering (ELC)“ beinhaltet ein kraftvolles Greifen mit der Großzehe, einen plantigraden (auf Sohlen gehenden) Fuß, gestreckte Hüfte und Knie, Beugung und Streckung sowie Einwärts- und Auswärtsdrehung des Ellenbogens in großem Umfang, ein bewegliches Handgelenk, gekrümmte Fingergrundglieder und ein stark gekrümmtes Daumengrundgelenk („deep first metacarpal joint“).
Danuvius unterscheidet sich nach Böhme et al. (2019) von bisher bekannten Affen in seiner Körperhaltung. Plantigraden und palmigraden quadrupeden1 Primaten (Altweltaffen und miozäner Menschenaffe Ekembo) fehlen suspensorische (hangelnde) Merkmale der Vorderextremitäten und die Streckmerkmale der Knie. Knöchelgehern (Schimpansen, Zwergschimpansen, Gorillas) fehlen gestreckte Knie, und die Fähigkeit zum Greifen von Großzehe und Daumen ist weniger kraftvoll entwickelt als bei Danuvius.
Den Fingergliedern von Danuvius fehlt die Robustheit, die typisch für Knöchelgeher ist.
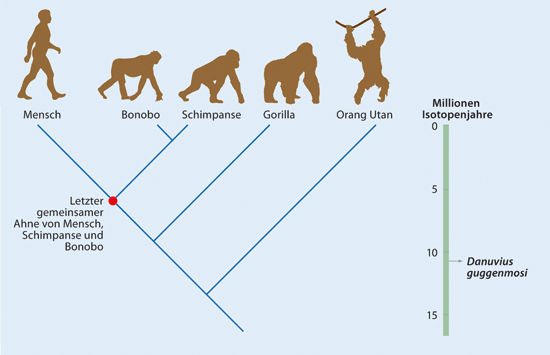
Abb. 3: Der evolutionäre Stammbaum von Mensch und Großaffen und die zeitliche Position des neuen Menschenaffen Danuvius guggenmosi. (Nach Kivell 2019)
Den in Bäumen herumturnenden Orang-Utans (Pongo) fehlen die gewichttragenden Anpassungen am Knie- und oberen Sprunggelenk und ihre Vordergliedmaßen sind stärker an die suspensorische Fortbewegung angepasst als bei Danuvius.
Weiterhin unterscheidet sich der neu entdeckte Menschenaffe von allen bisher bekannten Altweltaffen in der Wirbelsäulenmorphologie. Danuvius besitzt eine funktionell elongierte (verlängerte) Lendenwirbelsäule kombiniert mit einer Lordose*, welche dazu führt, dass die Körpermasse über die vergrößerten oberen Schienbeingelenkflächen bewegt wird.
Die Einmaligkeit beim ELC von Danuvius besteht nach Böhme et al. (2019) darin, dass nicht wie bei den meisten Primaten die Vorder- oder die Hinterextremität eine führende Rolle bei der Fortbewegung spielt, sondern dass beide in etwa gleichen Teilen daran beteiligt sind. Nach Böhme et al. (2019) sind bei Danuvius Gelenkpositionen und Belastungsmuster einer homininen Bipedie mit Hinterextremitätenstreckung und Wirbelsäulenkrümmung und eine suspensorische großaffenähnliche Fortbewegung mit kraftvollen mobilen Vorderextremitäten kombiniert. ELC repräsentiert nach Böhme et al. (2019) ein neues Modell der Körperhaltung des letzten gemeinsamen Ahnen der lebenden Großaffen und des Menschen (Abb. 2).
ELC, wie es Danuvius praktizierte, soll demnach ein Vorläufer der gewohnheitsmäßigen Bipedie gewesen sein.
Aus dieser Fortbewegung sollen sich dann die spezialisierteren Bewegungsformen entwickelt haben: Knöchelgang der Afrikanischen Großaffen (Gorilla, Schimpanse), das reine Schwingen des Orang-Utans und das gewohnheitsmäßige zweibeinige Schreiten des Menschen. Abb. 3 zeigt den evolutionären Stammbaum von Mensch und Großaffen und die zeitliche Position des neuen Menschenaffen Danuvius guggenmosi.
Im Folgenden werden die Deutungen der Knochenmerkmale von Danuvius im Rahmen der Fortbewegungsweise von Böhme et al. (2019) einer kritischen Betrachtung unterzogen.
Glossar
Abduktor: Muskel, der vom Körper wegführt. Artikulation: Gelenkverbindung. biped: zweibeinig. Condylus: Gelenkkopf eines Knochens. costal: zur Rippe gehörend. craniodental: zu den Zähnen und dem Schädel gehörend. Dorsalflexion: Beugung eines Bewegungssegments nach dorsal, d. h. zum Rücken, Handrücken oder Füßrücken hin. Glutealmuskeln: Muskeln, die das Gesäß formen. Homininen: Menschenähnliche; damit werden alle fossilen und lebenden Menschenformen einschließlich ihrer im Evolutionsmodell vermuteten Vorläufer bezeichnet. inferolateral: unten und seitlich. Inversion: Kombinationsbewegung aus Auswärtsdrehung, Heranführung an den Körper und Bewegung in Richtung der Sohlenrichtung des Fußes. Kyphose: Krümmung nach hinten. Lordose: Krümmung nach vorn. Malleolus: hammerförmiger Knochenvorsprung an Schienbein und Wadenbein. medioateral: zur Mitte und Seite hin. posterosuperior: hinten und oben. Processus transversus: paariger Querfortsatz eines Wirbels. trochlear: rollenförmig.
Kritische Beurteilung morphofunktioneller Schlüsse
Spekulative Schlüsse
Böhme et al. (2019) schließen von zahlreichen Merkmalen des Körperstamm- und Extremitätenskeletts von Danuvius auf die Fortbewegungsweise von Danuvius. Einige dieser Schlüsse sind aufgrund der bekannten Datenbasis berechtigt, andere müssen jedoch als hochspekulativ eingestuft werden.
Je ähnlicher Knochenmerkmale und insbesondere Knochenmerkmalskomplexe fossiler Formen denen bekannter lebender Wesen sind und je stringenter dabei morphofunktionelle* Zusammenhänge nachgewiesen sind, desto sicherer können funktionelle Schlüsse von Knochen bei fossilen Formen gezogen werden. Auf dieser Basis dürfte die Fortbewegung von Danuvius in Bäumen unstrittig sein. Darauf weist zum einen das Längenverhältnis der Elle zum Schienbein hin, welches ähnlich dem von Zwergschimpansen ist, und zum anderen die Morphologie der Elle, die einen großen Bewegungsumfang und die deutliche Krümmung der Fingergrundglieder nahelegt. Bei vielen anderen Merkmalen, insbesondere die unteren Extremitäten betreffend, sind die Schlüsse von Böhme et al. (2019) weit weniger sicher.
Zum einen sind die aufeinander aufbauenden knochenmorphologisch funktionellen Schlüsse von Böhme et al. (2019) fraglich, da sie spekulativer Natur sind, insbesondere weil wir es mit heute unbekannten Merkmalsmosaiken zu tun haben. Als Beispiele für solche Schlussketten seien der obere Teil des Oberschenkelknochens, der untere Teil des Schienbeines und die Wirbelsäule aus der Arbeit von Böhme et al. (2019) angeführt.
1. Proximaler Oberschenkelknochen: Von einem niedrigen Trochanter major, einem mehr vertikal ausgerichteten Hals und einer posterosuperioren* großen Kopfgelenkfläche des Femurs wird auf ein gewohnheitsmäßig gestrecktes Hüftgelenk, ein nach außen gedrehtes Hüftbein und eine Kippung der Beckenschaufel nach inferolateral* geschlossen. Von dieser Knochenkonfiguration leitet man dann Glutealmuskeln* ab, die als Abduktoren* die Hüfte bei einer bipeden Haltung wie bei den Homininen stabilisiert haben sollen.
2. Distales Schienbein: Von einem mediolateral* kurzen vorderen trochlearen* Rand und einem mediolateral schmalen Malleolus* wird auf eine mehr gleichmäßige Verteilung der auf die Gelenkfläche wirkenden Kräfte und eine geringe Belastung des oberen Sprunggelenkes in Dorsalflexion* und Inversion* im Vergleich mit den lebenden Großaffen geschlossen.
3. Wirbelsäule: (1) Von einer gut entwickelten Gelenkfläche für die Artikulation* mit der Rippe eines Brustwirbels wird auf eine Position des Zwerchfells oberhalb des untersten Thorakalwirbels geschlossen. Von dieser Zwerchfellposition wird auf einen funktionell längeren unteren Rücken, von diesem wiederum auf eine Lordose* der Lendenwirbelsäule und von dieser schließlich auf eine (menschenähnlichere) effektive Position des Masseschwerpunktes über den gestreckten Hüften, Knien und den plantigraden Füßen geschlossen. (2) Von der dorsalen Ausrichtung der Processus transversi* der Brustwirbel in Verbindung mit einem niedrigen Winkel der costalen* Gelenkflächenwinkel am ersten Brustwirbel wird auf einen mediolateralen Brustkorb und eine aufrechte Körperhaltung geschlossen. (3) Von der unterschiedlichen Neigung des Processus spinosus zwischen dem ersten und dem untersten Brustwirbel werden eine Lordose der Halswirbelsäule und eine Kyphose* der Brustwirbelsäule abgeleitet.
Wie spekulativ Vermutungen über Knochen-Funktionsbeziehungen gerade im Bereich der unteren Extremitäten sind, haben empirische Untersuchungen an zweibeinig gehenden Großaffen und Menschen eindrucksvoll gezeigt. Bisher ging man in der Forschung davon aus, dass der Mensch einen rigiden und die Großaffen einen mobilen Midfoot (vordere Fußwurzel) besitzen. Der rigide Midfoot wurde in Beziehung zu einem nur beim Menschen vorkommenden Längsfußgewölbe und als Voraussetzung für seinen einmaligen gewohnheitsmäßig schreitenden Gang auf zwei Beinen angesehen. Neuere Untersuchungen haben diese Vorstellungen völlig widerlegt.
Untersuchungen der Funktion
des Fußes lebender Großaffen und Menschen haben bisherige von der Knochenstruktur abgeleitete funktionelle Vorstellungen grundlegend revidiert.

Abb. 4: Bipedie des Orang-Utans (Pongo) und des Schimpansen (Pan) beim Klettern. A Mit den Armen unterstützte Bipedie mit gestreckten Knien beim Orang-Utan. B Mit den Armen unterstützte Bipedie mit gebeugten Knien beim Schimpansen. (Nach Thorpe et al. 2007, Abb. 1, A und D)
Das Längsfußgewölbe ist nicht wie bisher angenommen eine anatomisch-statische, sondern eine temporäre Struktur, die auch bei Großaffen auftritt. Damit einhergehend wurde gezeigt, dass nicht nur der Großaffen-Midfoot, sondern auch der menschliche Midfoot mobil ist, obwohl die Gelenkstruktur des Midfoot bei beiden sehr verschieden ist. Von der Gelenkmorphologie des Midfoot kann somit nicht auf seine Mobilität und von der Midfoot-Mobilität nicht auf die spezifische Kinematik des Fußes beim zweibeinigen Gehen geschlossen werden. Im Textkasten sind diese unerwarteten Untersuchungsergebnisse ausführlich dargestellt. Sie mahnen zu großer Vorsicht bei funktionellen Deutungen fossiler Knochenmerkmale, insbesondere bei Merkmalsmosaiken, für die es heute keine Analogie gibt.
Interessant in diesen Zusammenhängen und in Anbetracht der postulierten Bipedie mit gestreckten unteren Extremitäten von Danuvius ist die Tatsache, dass auch der Orang-Utan (Pongo) beim Stehen auf zwei Beinen in Bäumen in 90 % der Fälle seine Kniegelenke streckt. Allerdings stützt Pongo diese Haltung in 75 % der Fälle mit den Händen ab. Im Gegensatz zu Pongo sind beim Schimpansen in bipeder Haltung die Knie gebeugt (Thorpe et al. 2007; Abb. 4). Beim Orang-Utan ist bemerkenswerterweise der äußere Condylus* des Schienbeins wie beim Schimpansen konvex geformt, bei Danuvius ist er dagegen flach ausgebildet (Böhme et al. 2019). Von der Knochenstruktur kann man bei Pongo nicht auf die Fähigkeit zu einer bipeden aufrechten Körperhaltung mit gestreckten Knien schließen. Auch dies zeigt, dass spezifische Schlüsse von Knochenmerkmalen auf die Fortbewegung kaum möglich sind.
Keine Korrelation zwischen Knochenansatz und Belastung von Bändern
Eine weitere grundsätzliche Kritik an den knochenmorphologisch-funktionellen Schlüssen von Böhme et al. (2019) betrifft Aussagen bezüglich der Bandstrukturen. Nach Böhme et al. (2019) weist die außergewöhnlich stark ausgebildete Eminentia intercondylaris (Erhebung zwischen den beiden Condylen des Schienbeins) bei Danuvius auf stark entwickelte Kreuzbänder hin. Außerdem erwähnen die Autoren kräftige Knochenkämme für den Ansatz der bindegewebigen Scheiden der Sehnen der Beugemuskulatur an den Fingergrundgliedern als Hinweis auf eine suspensorische Fortbewegung. Dies impliziert den Schluss von starken Knochenansätzen auf dort ansetzende stark ausgebildete bindegewebige Scheiden und von diesen wiederum auf starke Handbeugemuskeln. An der Großzehe schließen Böhme et al. (2019) von gut entwickelten knöchernen Ansätzen auf gut entwickelte Muskeln.
Für diese Zusammenhänge gibt es keine Belege. Im Gegenteil, bisherige Untersuchungen konnten keinen Zusammenhang zwischen der Muskelstärke und der Größe und Struktur des Knochenansatzes der Muskelsehne (Enthese) nachweisen. Es existiert somit keine Korrelation zwischen der Belastung und der Ausbildung der Enthese, wie Böhme et al. (2019) in ihrer Argumentation voraussetzen (Zumwalt 2006, Rabey et al. 2015, Williams-Hatala et al. 2016, Wallace et al. 2017).
Böhme et al. (2019) vermuten weiter, dass die stark entwickelten Kreuzbänder Drehmomenten entgegenwirken, die durch Rotation des Körpers über die Knie entstehen. Sowohl die funktionelle Rolle der Kreuzbänder als auch die postulierte Rotation des Körpers über die Knie sind Spekulationen, ebenso die anderen oben angeführten knochenmorphologisch-funktionellen Kausalketten.
Midfoot-Beweglichkeit, Längsfußgewölbe und die Evolution der zweibeinigen Fortbewegung – ein paläanthropologisches Lehrstück
Nach klassischer Vorstellung besitzt der Mensch im Gegensatz zu allen nichtmenschlichen Primaten ein anatomisch-statisches Längsfußgewölbe. Die Existenz des Längsfußgewölbes wird als die wichtigste Anpassung des menschlichen Fußes an das zweibeinige Gehen angesehen und soll in Beziehung zu bestimmten Merkmalen der an ihm beteiligten Knochen stehen. Weiterhin glaubte man, dass das Längsfußgewölbe des Menschen eine Verminderung der Bewegung, also eine Versteifung der Gelenke des Midfoot (vordere Fußwurzel), bewirkt und so einen stabilen Hebel für das effektive Abstoßen des Fußes beim zweibeinigen Gehen liefert. Diese Vorstellung geht auf die beiden Autoren Elftman & Manter aus dem Jahr 1935(a) zurück, wurde später von Bojsen-Møller (1979) bestätigt und bis heute häufig zitiert.
Beim Schimpansen dagegen – so die Vermutung – ermöglichen die mobilen Midfoot-Gelenke ein effektives Greifen des Fußes bei der Fortbewegung in Bäumen. Auch diese Vorstellung geht auf die beiden Autoren Elftman & Manter ebenfalls aus dem Jahr 1935(b) zurück und wurde von anderen Autoren bestätigt (Susman 1983, DeSilva 2010) bzw. übernommen. Nach klassischer Vorstellung gibt es also eine funktionelle Dichotomie zwischen Großaffe und Mensch – mobiler versus rigider Midfoot.
Die größte Bedeutung beim zweibeinigen Gehen wird der letzten Phase, der Abstoßphase, zugemessen. Das Abstoßen des Fußes beginnt mit dem Abheben des Hackens vom Erdboden und endet, wenn die Zehen den Kontakt mit dem Erdboden verlieren. Während der menschliche Fuß nach dem Anheben des Hackens rigide (steif) bleibt, beugt sich der Fuß der Afrikanischen Großaffen leicht nach dorsal um die Midfoot-Gelenke (Elftman & Manter 1935b, D’Août et al. 2002, Vereecke et al. 2003, DeSilva 2010). Diese Bewegung um die Midfoot-Gelenke wird „midtarsaler Bruch“ genannt (Susman 1983).
Man suchte nach knöchernen Gelenkstrukturen, die zur Einschränkung der Midfoot-Beweglichkeit beim Menschen beitragen, und fand sie auch. Dazu gehören ein Verschlussmechanismus zwischen dem Fersenbein und dem Würfelbein (Bojsen-Møller 1979) und ein flaches Gelenk zwischen dem Würfelbein und dem Mittelfußknochen IV (De Silva 2010, Proctor 2013).
Bei den Afrikanischen Großaffen fehlen dagegen diese knöchernen Strukturen.
Man identifizierte außerdem Skelettmerkmale, welche mit der Existenz eines Fußgewölbes korrelieren (siehe Brandt 2017). Bezeichnenderweise besteht heute unter Paläanthropologen aber wenig Konsens darüber, welche Skelettmerkmale auf ein Längsfußgewölbe hinweisen (siehe Brandt 2017).
Bei fossilen Homininen wird anhand von Knochenmerkmalen auf den Beweglichkeitsgrad des Midfoot und auf die Ausprägung eines Längsfußgewölbes und damit auf den Entwicklungsgrad der zweibeinigen Fortbewegung auf der Evolutionsleiter zum Menschen hin geschlossen, z. B. bei Australopithecus afarensis (Ward et al. 2011) und „Homo“ habilis (OH 8) (Susman 1983).
Die jahrzehntelang angenommene Dichotomie in der Midfoot-Mobilität und damit einhergehende Existenz oder das Fehlen eines Längsfußgewölbes bei Mensch und Großaffe, die eine Schlüsselrolle in der Debatte bezüglich der Evolution der zweibeinigen Fortbewegung bei fossilen Homininen spielt, wurde aber durch neue empirische Studien widerlegt.
2008 stellten Lundgren et al. überraschend fest, dass beim Menschen beim zweibeinigen Gehen eine große Beweglichkeit im Bereich des äußeren Längsfußgewölbes besteht.
Die Ergebnisse einer Studie von Bates et al. (2013) lassen zudem erhebliche Zweifel an der Vorstellung aufkommen, dass die Midfoot-Mobilität und das äußere Längsfußgewölbe zu einem wesentlichen Teil knöchern bedingt und deshalb bei fossilen Hominiden erkennbar seien.
Bates und seine Kollegen ließen Menschen, die gewohnheitsmäßig Schuhe tragen, auf einem Laufband gehen und maßen die Druckverteilung im Bereich der Fußsohle.
Entgegen bisheriger Vorstellungen war die Druckverteilung im Bereich des äußeren Midfoot beim Menschen sowohl intrasubjektiv, also Schritt für Schritt bei einem Individuum, als auch zwischen den Individuen hochvariabel. Dabei wurde mit der Sohle des Midfoot auch der Boden berührt. Es trat also auch beim Menschen ein midtarsaler Bruch auf, der bisher als ein ausschließlich nichtmenschliches Phänomen angesehen wurde. Auch DeSilva & Gill (2013) haben dieses Phänomen beim Menschen nachgewiesen.
Aber auch zwei untersuchte Großaffen (Zwergschimpanse, Orang-Utan) zeigten wie der Mensch eine erhebliche Variation (Schritt-für-Schritt) des Druckes unter dem äußeren Midfoot. Die Druckverteilungen bei Menschen und bei Großaffen waren so variabel, dass sich die Werte sogar deutlich überlappten. Allerdings war der Mittelwert des Fußsohlendruckes des Menschen geringer als der der Großaffen. Nur dieses Ergebnis entsprach den Erwartungen, während alle anderen Ergebnisse bisherigen Vorstellungen widersprachen.
Die Studie von Bates et al. (2013) hat entgegen allen Erwartungen gezeigt, dass die Stabilität des äußeren Fußgewölbes beim Menschen nicht obligat, sondern oft nur vorübergehend und ein äußeres Längsfußgewölbe auch bei Großaffen nachweisbar und variabel ausgebildet ist. Das bedeutet jedoch nicht zwangsläufig, dass die Knochenstruktur im Bereich des äußeren Längsfußgewölbes gar nichts mit dieser offensichtlich nur temporär funktionellen Struktur zu tun hat.
Ein äußeres Längsfußgewölbe in Verbindung mit gewohnheitsmäßig praktizierter zweibeinig terrestrischer Fortbewegung kann nicht von den Fußknochenstrukturen abgeleitet werden. Dies könnte auch erklären, warum unter Paläanthropologen, wie schon oben erwähnt, wenig Konsens darüber besteht, welche Skelettmerkmale auf ein Längsfußgewölbe hinweisen. Bates et al. (2013, 6) schreiben zu dieser Thematik: „Das Versagen der Knochenmorphologie als unzweideutiger Hinweis auf die Fußfunktion in einem terrestrischen Zusammenhang erklärt vielleicht, warum kaum oder kein Konsens über das Vorhandensein eines medialen Längsfußgewölbes und/oder eines stabilisierenden lateralen Midfoot bei allen fossilen Homininen vor Homo erectus existiert …“
Greiner & Ball (2014) haben an Leichen des Menschen und Kadavern des Schimpansen überraschend festgestellt, dass deren Midfoot-Gelenke einen ähnlichen passiven Mobilitätsgrad besitzen. Allerdings ist dieser bei Makaken und Pavianen größer.
Holowaka et al. (2017) sind in ihrer Untersuchung noch einen wesentlichen Schritt über Greiner & Ball (2014) hinausgegangen. Die Autoren untersuchten die Bewegungen des Midfoot von Menschen und Schimpansen beim zweibeinigen Gehen während der Standphase in drei Ebenen. Die Forscher kamen zu dem Ergebnis, dass der Mensch nicht wie bisher vermutet einen geringeren, sondern sogar einen größeren Bewegungsbereich in den Gelenken des Midfoot beim zweibeinigen Gehen aufweist als der Schimpanse.
Allerdings weist der Schimpanse beim zweibeinigen Gehen währen der Abstützphase in der sagittalen Ebene eine größere Mobilität als der Mensch auf. Der Mobilitätsunterschied zwischen Mensch und Schimpanse in dieser Ebene ist aber viel geringer, als man vorher vermutet hatte.
Nicht primär die Gelenkmorphologie, sondern die Weichteilstrukturen mit ihren spezifischen Innervationen sind wahrscheinlich für die Versteifung des menschlichen Midfoot, die mit dem Anheben der Ferse stattfindet, verantwortlich, vermuten Holowka et al. (2017). Zusammenfassend ist festzustellen, dass der menschliche Fuß und der Großaffenfuß relativ mobil, aber auch zur Versteifung fähig sind. Die Kinematik des Fußes von Mensch und Großaffe beim bipeden Gehen unterscheidet sich. Die verschiedenen Weichteilstrukturen mit ihrer unterschiedlichen Innervation dürften zumindest teilweise für diese Unterschiede verantwortlich sein.
Entgegen jahrzehntelanger scheinbar gesicherter biomechanischer Vorstellungen mit dazu passenden knochenmorphologischen Befunden kann die Mobilität der Midfoot-Gelenke nicht von der Knochenstruktur erschlossen werden. Und auch wenn die Mobilität der Midfoot-Gelenke bekannt ist, kann von dieser nicht die spezifische Kinematik des Fußes beim zweibeinigen Gehen abgeleitet werden. Schon der Ausgangspunkt dieser Kette von nicht sicher möglichen Schlüssen, die tatsächliche Morphologie der Knochen, ist nicht selten Gegenstand kontroverser Diskussion.
Wenn eine Kette von Schlüssen mit der Knochenmorphologie als Ausgangspunkt und der spezifischen Kinematik der bipeden Fortbewegung am Ende selbst bei lebenden Primaten nicht möglich ist, dann gilt das umso mehr bei fossilen Primaten mit einem Knochenmerkmalsmosaik, für das es keine rezente Analogie gibt.
Bewertung von Danuvius
Böhme und ihre Kollegen (2019) haben mit Danuvius guggenmosi eine neue fossile Menschenaffenart entdeckt, die nach den craniodentalen Merkmalen aus dem Formenkreis der Dryopithecinen stammt. Der Affe aus dem bayerischen Allgäu wird auf 11,62 Millionen Jahre datiert. Die Körperproportionen von Danuvius sind ähnlich denen des Zwergschimpansen. Im Bereich des Körperstamm- und Extremitätenskeletts zeigt der neue Primat eine bisher unbekannte Kombination von Merkmalen, die auf eine unbekannte Fortbewegungsweise schließen lässt. Böhme et al. (2019) vermuten, dass sie eine Kombination von Suspension (Hangeln in Bäumen) und Bipedie mit gestreckten unteren Gliedmaßen war. Danuvius könnte nach Böhme et al. (2019) ein mögliches Modell der Fortbewegung des letzten gemeinsamen Ahnen von Großaffe und Mensch repräsentieren. Als realhistorisch letzter gemeinsamer Ahne von Schimpanse und Mensch ist Danuvius mit 11,6 Millionen Isotopenjahren zu alt, denn im heutigen Evolutionsmodell trennten sich Mensch und Schimpanse vor 5 bis 7 Millionen Isotopenjahren vom letzten gemeinsamen Ahnen.
Der Beginn der vermuteten Evolution des menschlichen zweibeinigen Gehens erfolgte nach Böhme et al. (2019) fortbewegend in Bäumen. Nach heutigem Kenntnisstand weisen einige Merkmale von Danuvius guggenmosi eindeutig auf Fortbewegungsaktivitäten in Bäumen hin. Weitere Schlüsse zur Fortbewegung des neuen Menschenaffen sind jedoch sehr unsicher, da die knochenmorphologisch-funktionellen Kettenschlüsse von Böhme et al. (2019) spekulativ sind.
Kivell (2019) weist in einem Begleitkommentar in Nature zu der Veröffentlichung von Böhme et al. (2019) darauf hin, dass viele fossile Großaffen aus dem Miozän einen Mix von Skelettanpassungen aufweisen, der unähnlich denen der lebenden Großaffen ist. Oft könne man nur raten, wie sich diese Großaffen fortbewegten und wie viel Zeit sie dabei in Bäumen oder auf dem Erdboden verbrachten. Diese Einschätzung dürfte im Grundsatz auch auf Danuvius zutreffen, da dessen Gesamtfortbewegungsrepertoire aufgrund des einmaligen Merkmalsmixes ebenfalls nicht sicher zu bestimmen ist.
Falls Danuvius sich tatsächlich biped mit gestreckten unteren Extremitäten in Bäumen fortbewegt hat, wie Böhme et al. (2019) vermuten, entstand gemäß Evolutionsmodell der zweibeinige Gang schon bei den Menschenaffen lange vor Abspaltung der Homininen vom gemeinsamen Großaffe-Mensch-Vorfahren und dies geschah auch nicht in Afrika, wie bisher postuliert, sondern in Europa. Die Bipedie wäre damit nicht mehr das entscheidende Identifikationskriterium von Homininen. Die im Evolutionsmodell als frühe Homininen klassifizierten Primaten mit Kletteranpassungen könnten deshalb auch Großaffen mit einem heute nicht mehr zu beobachtenden Fortbewegungsmix gewesen sein.
Im Grundtypkonzept der Schöpfungslehre repräsentiert Danuvius guggenmosi eine ausgestorbene Art eines Menschenaffen-Grundtyps. Innerhalb von Grundtypen sind aufgrund angelegter (latenter) genetischer Programme zahlreiche Skelettmerkmalskombinationen zu erwarten. Menschenaffen, die sich in Bäumen hangelnd und mit gestreckten Beinen biped auf Baumzweigen abstützend fortbewegten, sind in diesem Modell gut denkbar.
1 Bei der plantigraden vierfüßigen Fortbewegung wird die gesamte Fußfläche und bei der palmigraden vierfüßigen Fortbewegung die gesamte Handfläche auf die Unterlage aufgesetzt.
The evolution of compliance in the human lateral mid-foot. Proc. R. Soc. B 280, 20131818.
A new Miocene ape and locomotion in the ancestor of great apes and humans. Nature 575, 489–493.
Calcaneocuboid joint and stability of the longitudinal arch of the foot at high and low gear push off. J. Anat. 129, 165–176.
Wie sicher sind Deutungen in der Paläanthropologie? Australopithecus sediba und sein merkwürdiges Merkmalsmosaik. In: Brandt M: Frühe Homininen. Eine Bestandsaufnahme anhand fossiler und archäologischer Zeugnisse. Stud. Integr. Special 1, S. 7–48.
Segment and joint angles of hind limb during bipedal and quadrupedal walking of the bonobo (Pan paniscus). Am. J. Phys. Anthropol. 119, 37–51.
Revisiting the “midtarsal break”. Am. J. Phys. Anthropol. 141, 245–258.
Brief communication: a midtarsal (midfoot) break in the human foot. Am. J. Phys. Anthropol. 151, 495–499.
Chimpanzee and human feet in bipedal walking. Am. J. Phys. Anthropol. 20, 69–79.
The evolution of the human foot, with especial reference to the joints. J. Anat. 70, 56–67.
Kinematics of primate midfoot flexibility. Am. J. Phys. Anthropol. 155, 610–620.
Chimpanzee and human midfoot motion during bipedal walking and the evolution of the longitudinal arch of the foot. Journal of Human Evolution 104, 23–31.
Fossil ape hints at how bipedal walking evolved. Nature 575, 445–456.
Invasive in vivo measurement of rear-, mid- and forefoot motion during walking. Gait Posture 28, 93–100.
Proximal metatarsal articular surface shape and the evolution of a rigid lateral foot in hominins. J. Hum. Evol. 65, 761–769.
Locomotor activity influences muscle architecture and bone growth but not muscle attachment site morphology. J. Hum. Evol. 78, 91–102.
Evolution of the human foot: evidence from Plio-Pleistocene hominids. Foot Ankle 3, 365–376.
Origin of human bipedalism as an adaptation for locomotion on flexible branches. Science 316, 1328–1331.
Dynamic plantar pressure distribution during terrestrial locomotion of bonobos (Pan paniscus). Am. J. Phys. Anthropol. 120, 373–383.
Physical activity alters limb bone structure but not entheseal morphology. J. Hum. Evol. 107, 14–18.
Ward CV, Kimbel WH & Johanson DC (2011)
Complete fourth metatarsal and arches in the foot of Australopithecus afarensis. Science 331, 750–753.
Morphology of muscle attachment sites in the modern human hand does not reflect muscle architecture. Sci. Rep. 6:28353.
The effect of endurance exercise on the morphology of muscle attachment sites. J. Exp. Biol. 209, 444–454.
Themen | Kurzbeiträge | Streiflichter
Studiengemeinschaft WORT und WISSEN e.V.
Letzte Änderung: 6/25/21
Webmaster