
Themen • Kurzbeiträge • Streiflichter
Mendel‘sche Artbildung und die Entstehung der Arten
2. Latente Information und präexistente genetische Programme
1. Woher kommt die Artenvielfalt?
3. Fortpflanzungsisolation und adaptive Radiationen
von Nigel Crompton
Studium Integrale Journal
27. Jahrgang / Heft 1 - Mai 2020
Seite 12 - 19
Zusammenfassung: Es sind mehrere Mechanismen bekannt, durch die bereits angelegte Variationen und Variationsprogramme verborgen bleiben oder ausgeprägt werden können. Dieses präexistente Potenzial ermöglicht eine schlüssige Erklärung für die Entstehung der Arten innerhalb genetischer Familien (Grundtypen) – ohne Mutationen.
• • • • •
Einführung
Im ersten Teil unserer Serie wurde Mendels Gesetz über die Kombinationen von Merkmalen vorgestellt und gezeigt, dass latente (verborgene) genetische Informationen ein enormes Variationspotenzial beinhalten können. Wie aber werden die angelegten, schon vorhandenen (präexistenten) genetischen Variationsprogramme latent* gehalten und durch welche Mechanismen können sie abgerufen werden? Unterschiedliche Ausprägung dieser Information kann schließlich zu Artbildung* führen, also zur Bildung verschiedener Arten innerhalb einer genetischen Familie (Grundtyp).
Mit einem Stern* versehene Begriffe werden im Glossar erklärt.
Kompakt
Es sind mehrere Mechanismen bekannt, durch die bereits vorhandene Erbinformation latent gehalten werden kann, also nicht ausgeprägt wird. Als Ursachen dafür werden Dominanz, Epistasis und Transposition beschrieben und an Beispielen erläutert.
Dominante Gene können rezessive Gene im heterozygoten (mischerbigen) Zustand unterdrücken, die im reinerbigen (homozygoten) Zustand zur Ausprägung kommen können. Epistasis ist eine Form der Gen-Gen- (bzw. Protein-Protein-)Wechselwirkung und kann dazu führen, dass nicht nur einzelne Gene, sondern auch ganze Entwicklungsprogramme latent gehalten werden, wenn das hypostatische („unterdrückte“) Gen ein Regulationsgen ist. Transposition ist das Springen von Genen (Transposons) im Erbgut, das ebenfalls dazu führen kann, dass latente Geninformation zur Ausprügung gelangt; dieser Vorgang ist – anders als die Homozygotisierung – reversibel.
Der Verlust der Mischerbigkeit kann zur Bildung verschiedener Arten innerhalb genetischer Familien* führen. Ausgehend von einem panheterozygoten Vorfahr (der in sehr vielen Merkmalen mischerbig ist) sind unterschiedlichste Kombinationen homozygot gewordener Gene und entsprechender Merkmalszustände denkbar, was einen enormen Pool an Möglichkeiten für spezialisierte Arten bildet. Die genetische Information ist bereits vorhanden und deshalb sind keine Mutationen nötig.
Latente Information: Dominanz, Epistase und Transposition

Abb. 1: Hybriden (Mitte, F1) aus Königspudel und Wolf (oben, P) zeigen eindrucksvoll, dass Merkmalsausprägungen der Eltern im Hybriden verborgen sein können. Diese latenten Merkmalsausprägungen können dann in den Nachkommen der Hybriden (unteres Bild, F2) in verschiedensten Weisen zum Vorschein kommen. (Universität Kiel)
Wie wird die Information für die Merkmale im Erbgut latent gehalten, ohne ausgeprägt (exprimiert*) zu werden? Dass Information latent vorhanden sein kann, ist für Biologen nichts Ungewöhnliches. So ist in den Chromosomen einer Zygote* Information latent vorhanden, und sie reicht aus, um für die vielen Merkmale zu codieren, die zur Herstellung aller weiteren Zellarten eines ganzen Organismus notwendig sind. Beispielsweise ist in den Zellen eines Kakteenstocks Information für die schönsten Blüten latent vorhanden. Oder in den Zellen von Raupen ist Information für farbenfrohe Schmetterlinge latent enthalten. In den Zellen unseres eigenen Körpers ist Information für das andere Geschlecht latent vorhanden. In ähnlicher Weise ist die Information latent (nicht exprimiert) vorhanden, die für die vielen Merkmalsausprägungen codiert, welche für die Bildung der vielfältigen von einem Organismus abstammenden Arten erforderlich sind.
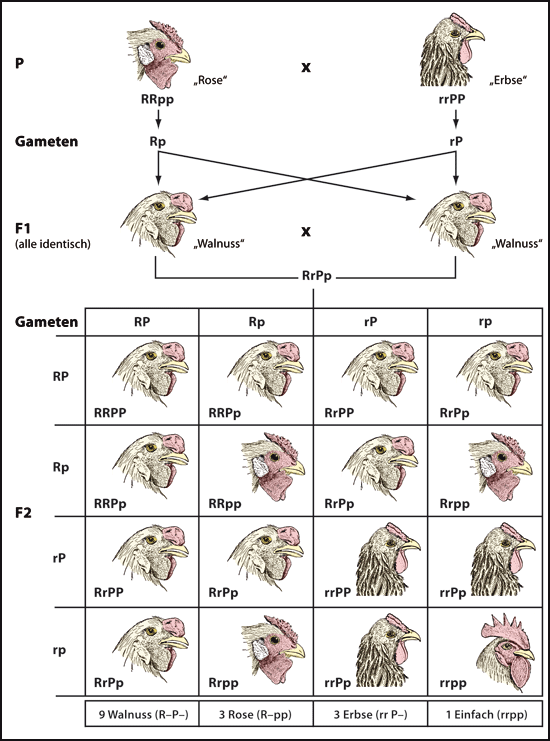
Abb. 2: Zwei Gene (jeweils mit zwei Allelen) wechselwirken miteinander, um vier Merkmalsausprägungen eines einzelnen Merkmals auszubilden – sogenannte Gen-Interaktion –; nach Studien der Genetiker Reginald Punnett und William Bateson von der Cambridge University. Gezeigt werden die Nachkommen der Selbstkreuzung von „Walnusskamm“, die wiederum durch eine Kreuzung von „Rosenkamm“ mit „Erbsenkamm“ entstanden ist. Hähne benutzen ihre Kämme zur Kühlung und zur Partnergewinnung. Hühner (bzw. Vögel im Allgemeinen) haben keine Schweißdrüsen und ihre Kämme (und Kehllappen) bieten eine große Oberfläche für den Wärmeaustausch. Die vier Hahnenkamm-Merkmalsausprägungen sind „Walnuss“, „Rose“, „Erbse“ und „Einfach“. Sie treten im vorhergesagten Mendel‘schen Verhältnis von 9:3:3:1 auf. Bei Kreuzungsversuchen hat man nie Hybriden mit nur „Walnuss“ und „Einfach“ in den Nachkommen beobachtet, deshalb wurde zum ersten Mal eine Gen-Interaktion vorgeschlagen (Bateson & Punnett 1905). Mittlerweile betrachtet man Merkmalsausprägungen als Konsequenz von Wechselwirkungen zwischen Genen als normal. „Einfach“ ist der bekannte und auffällige Hahnenkamm. Die unregelmäßigen „Rosen“- und „Erbsen“-Kämme sind beide dominant über „Einfach“. Sie gehen auf Mutationen zurück, die durch eine unzweckmäßige Überexpression der zwei Gene MRN2 und SOX5 im Kopf des Embryos verursacht werden. „Rose“ und „Erbse“ wirken zusammen und bilden den stark reduzierten „Walnuss“-Kamm (Imsland et al. 2012; Boije et al. 2012).
Es sind mindestens drei Mechanismen bekannt, wie die Information für die Merkmalsausprägungen in einem latenten Zustand gehalten werden: Dominanz, Epistasis und Transposition.
Dominanz
Dominanz ist die erste Möglichkeit, wie phänotypische Information in einem latenten, nichtexprimierten Zustand gehalten wird (Abb. 1). Von Mendel stammen die Begriffe „dominant“ und „rezessiv“. Er schrieb: „Der Ausdruck ,recessiv‘ wurde deshalb gewählt, weil die damit benannten Merkmale an den Hybriden* zurücktreten oder ganz verschwinden, jedoch unter den Nachkommen derselben, wie später gezeigt wird, wieder unverändert zum Vorschein kommen.“ Häufig ist der Unterschied zwischen einem dominanten Allel und einem rezessiven Allel nur eine Frage der Funktionalität. Ein dominantes Allel führt zu einem funktionsfähigen Protein, während ein rezessives Allel ein Protein mit beeinträchtigter Funktion oder vielleicht gar kein Protein zur Folge hat. Alternative Eigenschaften können jedoch viel komplexer sein (vgl. Abb. 2).
Epistasis
Epistasis (Abb. 3, 4) ist eine spezielle Form der Gen-Interaktion. Die Wechselwirkung von Genen (bzw. ihren Proteinen – auch Gen-Interaktion genannt) wurde zuerst bei den Ausprägungen des Kamms des Hahnes beobachtet und damals bezeichnet als „Koexistenz in einer Zygote von Elementen, die zu verschiedenen allelomorphen Paaren gehören“ (Bateson & Punnett 1905). Zwei Gene mit jeweils zwei Allelen bringen vier Merkmalsausprägungen eines Merkmals hervor (Abb. 2). In diesem Fall handelt es sich allerdings nicht um Epistasis. Epistasis liegt dann vor, wenn zumindest eine der vier Merkmalsausprägungen fehlt. Sie unterscheidet sich von der Dominanz, bei der die dominante Merkmalsausprägung die rezessive Merkmalsausprägung einfach verdrängt. Bei Epistasis dagegen verbirgt das Vorhandensein einer Merkmalsausprägung eines Gens (bzw. eines Allels*), das man epistatisch nennt, die Merkmalsausprägungen eines anderen Gens, das man hypostatisch nennt. Hier handelt es sich immer noch um Mendel’sche Genetik und es gibt enstprechend dominante und rezessive Epistase.
Dominante Epistasis liegt vor, wenn ein einzelnes dominantes Allel eines Gens die Expression* der Allele des hypostatischen Gens unterdrückt. Als klassisches Beispiel gelten hier die Fruchtfarben des Kürbisses mit den Merkmalen weiß, gelb oder grün. Bei dieser Situation, bei der die Merkmalsausprägung der Farbe durch zwei Gene bestimmt wird, würde man vier Merkmalsausprägungen erwarten; da aber nur drei beobachtet werden, weist das darauf hin, dass hier Epistasis stattfindet. Das Vorliegen nur eines einzelnen dominanten epistatischen Allels bewirkt, dass nur weiße Früchte ausgebildet werden. Wenn das dominante epistatische Allel homozygot* (und damit fixiert) ist, werden nur weiße Früchte bei sich selbst kreuzenden Nachkommen beobachtet, gelbe und grüne Früchte werden nie produziert (die Ausprägung dieser Farben wird unterdrückt). Dennoch kann die Information für gelbe und grüne Früchte durchaus latent im Erbgut vorhanden sein und in Hybriden exprimiert werden.
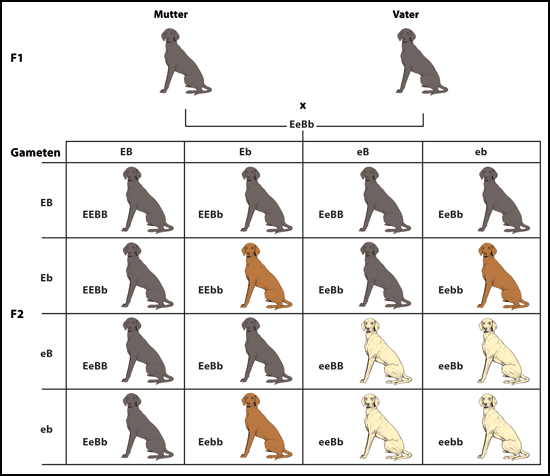
Abb. 3: Rezessive Epistasis am Beispiel der Fellfarbe dunkel (schwarz oder braun) und hell (goldfarben) beim Labrador. Es liegen zwei Gene zugrunde: B (B, b) und E (E, e). B ist dominant über b. Wenn wenigstens ein B vorliegt, ist das Fell schwarz, wenn b doppelt rezessiv ist, ist das Fell braun (BB, bB, Bb schwarz, bb braun).
Das Gen E entscheidet, ob das Fell dunkel oder hell ist. Liegt wenigstens ein E vor, ist das Fell dunkel, ist e doppelt rezessiv, dann ist das Fell hell (goldfarben). Die helle Farbe entsteht dadurch, dass nur Phaeomelanin gebildet wird. Zur Ausprägung der dunklen Farbe muss Eumelanin produziert werden. In goldfarbenen Labradoren sind die Merkmalsausprägungen schwarz und braun vorhanden, bleiben aber latent.
Das Allel B / b entscheidet also, welche der beiden dunklen Ausprägungen realisiert wird; das Allel E / e entscheidet, ob überhaupt dunkles Pigment (Eumelanin) produziert wird (nämlich dann, wenn wenigstens eines der Allele E ist).
E ist somit rezessiv epistatisch über B, d. h. E kann B „ausschalten“, wenn es doppelt rezessiv vorliegt (ee); B verbleibt dann aber latent im Erbgut.
Rezessive Epistasis liegt vor, wenn beide Allele des epistatischen Gens in rezessiver Form vorhanden sein müssen, um die Ausprägung des hypostatischen Gens zu unterdrücken. Ein klassisches Beispiel dafür ist die Fellfarbe des Labradors mit den Merkmalen gold, schwarz oder braun (Abb. 3). Auch hier erwartet man mit zwei Genen vier Merkmalsausprägungen, und weil man nur drei sehen kann, weist dies darauf hin, dass Epistasis stattfindet. Wenn beide Allele des epistatischen Gens in rezessiver Form vorliegen, kommt nur die goldene Fellfarbe vor. Schwarze und braune Labradore werden in diesem Fall bei selbst kreuzenden Nachkommen nie zu sehen sein. Dennoch ist die Information für schwarze und braune Labradore latent im Erbgut vorhanden und kann in Hybriden exprimiert werden.
Warum ist die Epistasis für die Mendel’sche Artbildung so wichtig? Bei einfacher Dominanz ist eine der beiden Merkmalsausprägungen typischerweise nichtfunktional (z. B. geht die Farbe verloren) und tritt in rezessivem Zustand nicht in Erscheinung. Wenn jedoch eine Epistasis vorliegt, können auch komplette Entwicklungsprogramme latent gehalten werden, sofern das hypostatische Gen ein Regulationsgen ist, und die Aufhebung der Epistasis aktiviert diese dann.
Ein Beispiel: Arabidopsis (Schmalwand) ist eine kleine krautige Pflanze aus der Familie der Kreuzblütler. Wenn zwei wichtige epistatische Suppressor-Gene ausgeschaltet werden, können sich ihre Nachkommen in einen Strauch wie eine Azalee verwandeln (Melzer et al. 2008; Abb. 5). Die Information für die Merkmalsausprägung ist zwar zuvor schon vorhanden, aber sie wird latent gehalten und wartet auf ihre Expression* durch Aufhebung der Epistasis. Von etwa 150 weiteren Kreuzblütlern in Mitteleuropa hat nur eine einzige Art, der im Jura-Gebirge vorkommende Felsen-Bauernsenf (Iberis saxatilis), einen holzigen Phänotyp. Eine Invasion von Inseln durch Arten krautiger Pflanzenfamilien wie Kreuzblütler, aber auch anderer Familien, führt jedoch zu einem unerwarteten Auftreten von vielen neuartigen holzigen Phänotypen (Arten und Gattungen) und auch anderen latenten Phänotypen wie latente Blütenfarben, Blütenstände oder Ausprägungen des Habitus (Carlquist 1971; 1974). Das Verholzungs-Programm und auch andere Programme sind also latent vorhanden und werden exprimiert, wenn die epistatische Unterdrückung ausgeschaltet wird.

Abb. 4: Epistasis kann auch doppelt auftreten, wie das Beispiel der Löffelenten zeigt, bei denen doppelte rezessive Epistasis (bzw. komplementäre Genwirkung) vorkommt. Die europäische Löffelente (Spatula clypeata, oben) und die Zimtente (S. cyanoptera, Mitte links) sind klar abgegrenzte eigenständige Arten. Interessanterweise sehen ihre Hybriden (unten) aus wie eine dritte Art, die australische Löffelente (S. rhynchotis, Mitte rechts). Besonders interessant ist der auffällige vertikale weiße Augenstreifen der Hybriden, der bei keiner der beiden Elternarten ausgebildet ist, was Folge einer doppelten Epistasis ist. Die Information für die Ausprägung des Augenstreifens ist also in den Elternarten vorhanden, dort jedoch latent. Im Hybriden dagegen wird sie ausgeprägt. (Aus Scherer & Hilsberg 1982)
Transposition
Von Transposition (Abb. 6) spricht man, wenn sich genetische Elemente (kurze DNA-Abschnitte) von einer Stelle in den Chromosomen zu einer anderen versetzen. Solche genetischen Elemente springen von einem Ort zum anderen. Je nachdem, wie sie sich versetzen, werden sie entweder Transposons oder Retrotransposons genannt. Man hat herausgefunden, dass diese Versetzungen die Ursachen für bestimmte Merkmalsausprägungen sind. Zwar konnte Mendel diesen Mechanismus oder dessen Auswirkungen noch nicht kennen, aber die Transposition spielt eine einzigartige, zentrale Rolle bei der Erhaltung der Vielfalt. Ihr ist mindestens eines der von Mendel untersuchten Merkmale zu verdanken (die Samenform, siehe Textkasten).
Der Unterschied zwischen rotem und weißem Wein wird zum Beispiel durch ein Retrotransposon namens Gret1 bewirkt, das in der Nähe eines Gens eingefügt wird, welches für die Regelung der Herstellung des roten Anthocyan-Pigments in Weintrauben zuständig ist. Gret1 blockiert die Expression dieses Gens, welches für den Transkriptionsfaktor VvmybA1 codiert. Wenn das Gen durch das Retrotransposon ausgeschaltet ist, wird kein Anthocyan mehr hergestellt und die Weintrauben werden nicht rot. Da nur eine funktionale Kopie des VvmybA1 für die Synthese von Anthocyan erforderlich ist, müssen beide Kopien des Gens ausgeschaltet werden, was z. B. eine Selbstkreuzung erforderlich macht, um die homozygote Form mit ihren zwei Kopien von VvmybA1 mit ausgeschaltetem Gret1 zu bilden. Das ist bei weißen Weinsorten der Fall (Abb. 7). Wenn Gret1 sich aber entfernt (herausspringt), erlangt das VvmybA1-Allel seine Aktivität zurück und es werden wieder rotes Pigment und rote Weintrauben erzeugt (Kobayashi et al. 2004).

Abb. 5: Mitte: Polsterartige Formen des Pflanzenwuchses einer soc1-3- und ful-2-Mutante von Arabidopsis thaliana nach 8 Monaten Wachstum unter Kurztagbedingungen. Links: Habitus der nicht mutierten Wildpflanze nach wenigen kurzen Tagen. Rechts: Verholzung bei Arabidopsis thaliana. (Aus: Melzer et al. 2008, Abdruck mit freundlicher Genehmigung)
Ein weiteres bemerkenswertes Beispiel für eine durch Transposition verursachte Merkmalsausprägung ist der Industriemelanismus bei Birkenspannern (Abb. 8), der sich aus der Einfügung eines großen Transposons in das erste Intron des cortex-Gens ergibt. In diesem Fall erzeugt das Transposon eine Verstärkung der cortex-Expression und dadurch eine dunklere Variante des Birkenspanners (van’t Hof et al. 2016).
Transposons und Mendel’sche Artbildung
Warum sind Transposons für die Mendel’sche Artbildung wichtig? Im Gegensatz zu alternativen Merkmalsausprägungen, die sich aus dem irreversiblen Verlust genetischer Informationen (durch Homozygotisierung) ergeben, können Transposons kommen und gehen oder sogar eine Feinregulierung der Genexpression bewirken. In diesen Fällen geht die Merkmalsinformation nicht verloren, wenn Gene homozygot „fixiert“ werden. Wenn sich die Transposons von einer bestimmten Position entfernen, wird die zuvor durch die Anwesenheit des Transposons unterdrückte Information wieder abgerufen. Die Transposition ermöglicht die vorübergehende Unterdrückung von genetischer Information für eine oder mehrere Generationen ebenso wie die darauffolgende erneute Freigabe.
Die drei Mechanismen Dominanz, Epistasis und Transposition wirken zusammen, um alternative Merkmalsausprägungen zu erzeugen. Rezessive Merkmalsausprägungen können aus einer einfachen Inaktivierung dominanter Funktionen resultieren, wie z. B. durch Basenpaar-Substitution, d. h. durch Mutation. Weitaus flexibler ist der Einsatz von eingefügten genetischen Elementen (z. B. Transposons), die in der Lage sind, Genexpression auf eine fein dosierte Weise zu regulieren. Umfangreiche wissenschaftliche Forschung zeigt, wie diese genetischen Elemente die spezifische Stärke einer bestimmten Funktion (oder eines Merkmals) regulieren können, ebenso die spezifische Position in Organen und Geweben, wo eine Funktion aktiviert ist, und sogar, unter welchen spezifischen Umweltbedingungen die Expression stattfindet. Solche genetische Elemente sind nicht für die Ausbildung der Merkmale selbst erforderlich, können aber elegant ihre alternativen Ausprägungen ermöglichen.
Glossar
Allel: Variante ein und desselben Gens. Artbildung: Aufspaltung einer Art (Spezies) in zwei Tochterarten. eukaryotisch: mit echten Zellkernen und damit zur Meiose fähig. Expression/exprimieren: Das Ablesen und somit Nutzbarmachen von Genen. exprimieren: → Expression. genetische Familie: Familie, deren zugehörende Arten durch verschiedene Merkmalskombinationen einer präexistenten Vielfalt charakterisierbar sind. heterozygot: mischerbig (das Gen liegt in zwei verschiedenen → Allelen vor.) homozygot: gleicherbig (das Gen liegt in zwei gleichen Allelen vor.) Hybrid: Mischling. Kooption: Übernahme von Genen für eine weitere Funktion. latent: versteckt, verborgen. Meiose: Reifeteilung, Bildung der Geschlechtszellen. Mutation: Spontane oder künstlich ausgelöste Änderung des Erbguts. Phänotyp: äußere Erscheinungsform. Spleißstelle: Position in einem Gen, an dem Teile des Gens vor der Bildung der mRNA herausgeschnitten werden. Transkriptionsfaktor: Regulationsprotein, das für die Initiation der RNA-Polymerase bei der Transkription (Übersetzung von DNA in mRNA) von Bedeutung ist. Zygote: Zelle von → Eukaryoten, die bei der geschlechtlichen Fortpflanzung durch Verschmelzung von einer Eizelle und einem Spermium entsteht.
Arten und genetische Familien
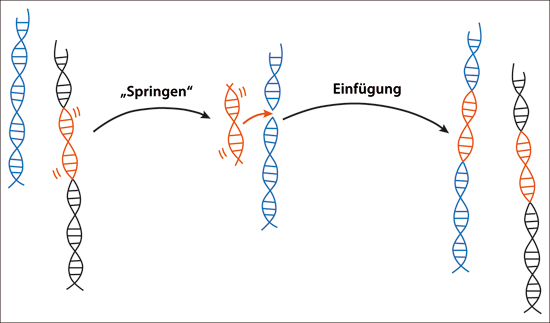
Abb. 6: Schematische Darstellung der Transposition eines Transposons. Dargestellt sind zwei verschiedene Chromosomen (blau und schwarz) mit einem Transposon (rot, links). Dieses Transposon kann sich entweder herausschneiden oder eine Kopie von sich produzieren. Beim „Springen“ befindet sich das Transposon neben dem blauen Chromosom (Mitte) und kann sich in dieses Chromosom einfügen („Einfügung” in das zweite (blaue) Chromosom, rechts). Beide Chromosomen erhalten ein Transposon nach dieser Form der Transposition.
In der Geschichte der Biologie wurden viele Definitionen des Artbegriffs vorgeschlagen. Manche davon basieren auf der Genetik. Wenn jedoch (vor allem in der Feldforschung) für praktische Untersuchungen auf diesem Gebiet etwas „Handfestes“ benötigt wird, werden Merkmalskombinationen für die Artdefinition zugrunde gelegt. Dabei wird eine Art – mehr oder weniger – durch einen Satz von Merkmalsausprägungen festgelegt. Ein anderer Satz von Merkmalsausprägungen definiert eine andere Art.
Für die Definition einer Art gelten im Allgemeinen einzelne Merkmalsausprägungen nicht als ausreichend. Mendel schrieb, dass sich gute Arten durch viele Merkmalsausprägungen unterscheiden: „Für die Versuche dienten grösstentheils Pflanzen, welche als gute Arten gelten und in einer grösseren Anzahl von Merkmalen verschieden sind.“ Er räumte allerdings ein, dass einige Erbsenvarianten als unabhängige Arten eingestuft werden könnten. Mendel bemerkte: „So wenig man eine scharfe Unterscheidungslinie zwischen Hybriden und Varietäten zu ziehen vermag, ebenso wenig ist es bis jetzt gelungen, eine klare Grenze zwischen den Species selbst und ihren Varietäten aufzustellen.“ Wenn man bedenkt, dass eine große Anzahl von Merkmalsausprägungen eine gute Art von der nächsten unterscheidet, stellt sich die Frage, was eine Art überhaupt zu einer Art macht. Die berühmten Silberschwerter von Hawaii gehören zu einem Pflanzenverband von drei Gattungen und 30 Arten. Diese alle sind so eng miteinander verwandt, dass sie leicht Hybriden untereinander bilden. Alle zusammen gehören offenbar zu einer genetischen Familie, was sich in der Kreuzbarkeit der zugehörigen Arten zeigt. Doch die Arten innerhalb genetischer Familien unterscheiden sich durch viele Merkmalsausprägungen und weisen außergewöhnlich unterschiedliche Pflanzenformen auf. Diese Situation ist für viele eukaryotische Arten typisch.

Abb. 7: Rote und weiße Traubensorten sind auf die Wirkung von Retrotransposons zurückzuführen (siehe Text). Links: U.S. Department of Agriculture – Agricultural Research Service (Gemeinfrei); Bild rechts: Kuschti (Gemeinfrei)
Das lässt die Frage aufkommen, welcher Mechanismus der Artbildung für die Entstehung einer so großen Anzahl von unterschiedlichen Merkmalsausprägungen innerhalb genetischer Familien verantwortlich sein kann. Ganz bestimmt reichen dafür einfache Mutationen nicht aus (s. u.). Hier bietet die Mendel’sche Artbildung, eigentlich seine Entdeckung der meiotischen Rekombination, eine völlig überzeugende Lösung. Die Meiose* beeinflusst gleichzeitig alle Gene in einem Organismus und kann deshalb leicht innerhalb einer einzigen Generation umfassende Änderungen in einem Phänotyp* des Organismus bewirken.
Verlust der Mischerbigkeit
Hybridisierung führt zur Heterozygotie (Mischerbigkeit). Umgekehrt führt Fortpflanzungsisolation dazu, dass die Heterozygotie verloren geht (Abb. 9). Der Verlust der Mischerbigkeit führt allmählich dazu, dass neue differenzierende (heterozygot-hybride) Merkmalskombinationen zu neuartigen konstanten (homozygot-reinerbigen) Merkmalskombinationen werden. Mendels Gesetz über die Merkmalskombinationen bedeutet, dass n einzigartige Merkmale 2n verschiedene Merkmalskombinationen hervorbringen können, von denen nur eine mit der ursprünglichen Elternhybride identisch ist (vgl. Teil 1 der Artikelserie). Die Nachkommen behalten dieselbe Anzahl von Merkmalen (Genen) bei. Es sind die Kombinationen der dominanten und rezessiven Merkmalsausprägungen (Allele), die sich unterscheiden und zu neuen Arten führen. Wenn die Mischerbigkeit verloren geht, werden mit jeder Generation die vorherigen differenzierenden (heterozygoten*) Merkmale immer konstanter (homozygoter). Wenn in den Nachkommen einer ursprünglichen Hybride eine geeignete (Art-definierende) Mischung von konstanten (reinerbigen) Merkmalen entsteht, haben diese sich zu neuen Arten verwandelt. Verlust der Mischerbigkeit lässt Hybriden zu verschiedenen Arten werden.
Artbildung: Mutationen und präexistente genetische Information
Eine umfangreiche phänotypische Veränderung ist eine natürliche Folge der Meiose und der Mendel’schen Artbildung durch zunehmende Homozygotie (vgl. Abb. 9). Der alternative Mechanismus, die Entstehung der Arten zu erklären, dem in genetischen und biologischen Lehrbüchern die uneingeschränkte Deutungshoheit zugestanden wird, ist die Mutation*. Mutationen werden als die einzigen Quellen genetischer Information angesehen. Doch einzelne Mutationsereignisse können weitreichende phänotypische Änderungen alleine nicht erklären. Zahlreiche vorteilhafte Mutationen könnten möglicherweise tiefgreifende Veränderungen mit sich bringen. Allerdings sind die bisher beobachteten Beispiele von Ereignissen mit vielfachen Mutationen stets schädlich für die Organismen.1 Das war schon immer die Achillesferse des Mutationsgeschehens. Mutationen sind ganz überwiegend mit Funktionsverlusten verbunden.

Abb. 8: Die helle und die dunkle Form des Birkenspanners (Biston betularia) sind auf die Wirkung von Retrotransposons zurückzuführen (siehe Text).
Allgemein werden immer wieder folgende zwei Beobachtungen gemacht: Mutationen bilden keine neue phänotypischen Merkmale und Mutationen bilden keine neue Arten (d. h. die nach Mendel „guten Arten“, s. o.). Vielfache nützliche Mutationen wären nötig, um auch nur ein einziges neues vorteilhaftes Merkmal zu erzeugen. Mehrfache Mutationen bringen jedoch erhebliche genetische und phänotypische Kollateralschäden mit sich. Denn Mutationen, die in latenter (rezessiver) Merkmalsausprägung als genetische Varianten akkumulieren, sind eine Quelle letztlich tödlicher Veränderungen. Akkumulierende Mutationsereignisse verschlechtern tendenziell genetische Information (Schoen et al. 1998; Uchimura et al. 2017). Mutation ist in erster Linie eine Folge der Entropie in der biologischen Welt, die unerbittlich biologische Information beseitigt, insbesondere latente Information in Eukaryoten.
Prozesse, die zur Artbildung beitragen
Es gibt drei Prozesse, die zur Artbildung beitragen. Der erste Prozess, die meiotische Rekombination vorhandener Merkmalsinformation, führt zur Entstehung vielfacher neuer phänotypischer Merkmalskombinationen. Der zweite Prozess, die Fortpflanzungsisolation, ist notwendig, um die Hybridisierung zu vermeiden, die zur Heterozygotie zurückkehren und damit die phänotypische Vielfalt rückgängig machen würde, welche durch die meiotische Rekombination gewonnen wurde, und die bewirkt, dass ausgeprägte Merkmale wieder latent werden (vgl. Abb. 9). Der dritte Prozess, die Selektion, „erhält begünstigte Rassen“ (Darwin 1859).
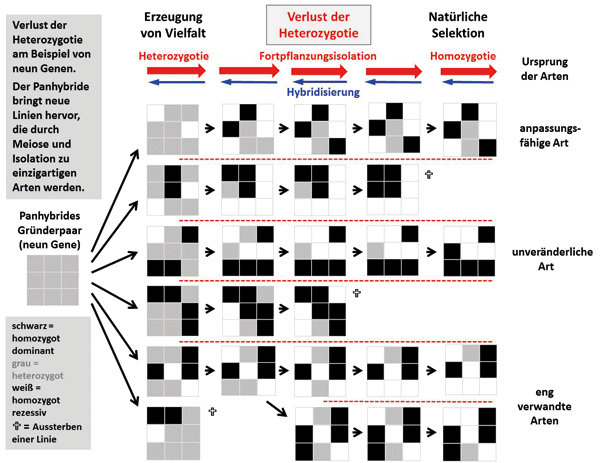
Abb. 9: Evolution eines Grundtyps (genetische Familie) mit neun Merkmalen. Genotypen der Population (bzw. Art) werden als 3x3-Matrizen dargestellt. Jede Spalte mit 3x3-Matrizen repräsentiert eine aufeinanderfolgende Generation und jede Zeile die evolutionäre Linie einer Population (bzw. Art). Jede Zelle in einer Matrix repräsentiert ein Merkmal: schwarz, homozygot dominant; grau, heterozygot; weiß, homozygot rezessiv.
Nachdem die Populationen isoliert wurden (gestrichelte Linien), variieren nur heterozygote Merkmale (grau). Der Verlust der Heterozygotie führt zu einzigartigen Konstellationen homozygoter dominanter und rezessiver Merkmale, d. h. zur Entstehung von Arten. Populationen (Arten) sterben aus verschiedenen Gründen aus (Kreuzsymbol). Die letzte Spalte stellt die heutigen Populationen (Arten) dar. Die meisten haben eine gewisse Heterozygotie und das Potenzial zur Diversifizierung bewahrt. Eine Art (3. Reihe) ist völlig homozygot und „unveränderlich“. Die Hybridisierung kann den Verlust der Heterozygotie (Pfeilkette) teilweise umkehren. In den beiden untersten Linien ist ein Verzweigungsereignis aufgetreten und die vorhandenen Nachkommen sind eng miteinander verwandt.
Der erste Prozess (meiotische Rekombination), der von Mendel (1866) entdeckt wurde, führt zur Ausprägung latenter phänotypischer Merkmalsausprägungen. Mendels Untersuchungen an Hybriden wurden an nahe miteinander verwandten Pflanzen durchgeführt, die sich in den Allelen eines, zweier oder dreier Gene unterschieden. Eine Kreuzung kann jedoch auch zwischen Individuen verschiedener Gattungen erfolgen und diese können sich in den Allelen von Tausenden von Genen unterscheiden. Arten innerhalb einer genetischen Familie teilen gemeinsame kompatible Gene, und Kreuzungen kommen natürlicherweise vor, wenn sie auch gewöhnlich unterdrückt werden. Arten aus separaten genetischen Familien besitzen jedoch inkompatible Garnituren von Genen; daher entstehen bei solchen Kreuzungen in der freien Natur keine Hybriden.
Der zweite Prozess, die Fortpflanzungsisolation, ist für die Erhaltung neuer Merkmalskombinationen sehr wichtig, welche aus neuartigen Kombinationen homozygoter dominanter und rezessiver Allele entstehen (siehe dazu die dritte Folge dieser Artikelserie).
Der dritte Prozess, die Selektion, die von Darwin (1859) so geschätzt und berühmt wurde, geschieht zwangsläufig. Selektion kommt in verschiedenen Formen vor: als natürliche, künstliche und sexuelle Selektion. Für die Entstehung neuartiger phänotypischer Merkmalsausprägungen bietet die Selektion allerdings keine Erklärung – sie beseitigt vorhandene Variation, erschafft sie aber nicht. Deshalb erklärt Selektion nicht die Entstehung der Arten. Sobald jedoch neuartige Merkmalsausprägungen aufgetreten sind, wird die Selektion, wenn diese Merkmalsausprägungen vorteilhaft sind, dafür sorgen, dass sie im Vergleich zu weniger vorteilhaften Merkmalsausprägungen innerhalb einer Population häufiger vorkommen. Falls die Merkmalsausprägungen aber unvorteilhaft oder weniger vorteilhaft als andere sind, wirkt die Selektion umgekehrt und entfernt diese Merkmalsausprägungen aus der jeweiligen Gruppe.
Latente genetische Information als entscheidende Quelle
Latente genetische Informati=on (Mendels rezessive Elemente) bleibt die alles entscheidende Quelle für phänotypische Neuheiten. Die Wissenschaftlergemeinschaft erkennt dies immer mehr an. In Fällen, bei denen Merkmale sogenannte nichtreduzierbare Komplexität aufweisen, muss eine Anzahl von funktionierenden Teilen gleichzeitig vorliegen und miteinander in Wechselwirkung stehen, damit das Ganze funktioniert. Es ist offensichtlich, dass Mutationsereignisse überhaupt nicht zufriedenstellend erklären können, wie solche nichtreduzierbar komplexen Merkmale entstanden sein könnten. In solchen Fällen wird Kooption* als Erklärung vorgebracht; man kann das als eine Art Neuverwendung von vorhandener Information bezeichnen. Der Grund für die Erklärung durch Kooption besteht darin, dass notwendige Information bereits vorhanden ist und somit problematische Szenarien mit unmöglichen Anzahlen von abgestimmten vorteilhaften Mutationsereignissen zum Erwerb neuer Information vermieden werden. Dieses Konzept läuft darauf hinaus, dass präexistente genetische Programme Artbildung möglich machen. So konnte z. B. gezeigt werden, dass die außergewöhnliche Radiation der Buntbarsche der großen afrikanischen Seen aufgrund der schon vorhandenen Variation möglich wurde (Brawand et al. 2014). In anderen Worten, bei Tausenden von Artbildungsereignissen wurde auf präexistente genetische Programme zurückgegriffen, und diese waren von Anfang an im Erbgut der Buntbarsche vorhanden.
Präexistente Variationsprogramme erlauben also eine schlüssige Erklärung für die Ausprägung vielfältiger Merkmale und sind eine wichtige Voraussetzung für Artbildung innerhalb genetischer Familien. Wie aber können entstehende Arten voneinander isoliert werden? Das ist Thema des dritten Teils dieser Artikelserie.
Hinweise: Eine ausführlichere Version dieses Artikels ist als mittlerer Teil eines Internetartikels veröffentlicht unter: https://www.wort-und-wissen.org/wp-content/uploads/b-19-3_mendel.pdf
Mendels Versuchsergebnisse aus heutiger Sicht
Dominanz, Epistasis und Transposition waren Mendel nicht bekannt. Dennoch schrieb er über die „innere Beschaffenheit“ der Hybriden (ihren Genotyp), und was weitervererbt wurde, nannte er „Elemente“, also das, was wir heute als Gene oder Allele bezeichnen. Er schrieb, dass die Hybridisierung bei der Befruchtung „in der materiellen Beschaffenheit und Anordnung der Elemente begründet ist, die in der Zelle zur lebensfähigen Vereinigung gelangten“.
Was wissen wir heute über die materielle Beschaffenheit und Anordnung der von Mendel so bezeichneten „Elemente“, der alternativen Allele? Mendel untersuchte sieben Erbsenmerkmale. Das erste Merkmal war die Samenform. Reife Samen haben entweder eine runde oder runzelige Form. Runzelige Erbsen sind kommerziell interessanter, weil sie süßer sind als runde Erbsen. Die runzelige Form ist rezessiv (r/r). Sie wird von einem Transposon verursacht, das in das Gen eingefügt wird, welches das stärkespaltende Enzym codiert. Diese Einfügung führt zu einer Anreicherung einfacher Zucker, was schließlich zu einem stärkeren Schrumpfen der Keimblätter und zu einer runzeligen Erbse führt (Bhattacharyya et al. 1990; Ried & Ross 2011).
Das zweite Merkmal war die Samenfarbe. Die Samen sind entweder gelb oder grün. Die grüne Form ist rezessiv (i/i). Sie ergibt sich aus dem Einfügen von sechs Nukleotiden, wodurch das sgr-Gen ausgeschaltet wird, das für die mit der Seneszenz verbundene Spaltung der Chlorophyll-Moleküle zuständig ist (Armstead et al. 2007; Sato et al. 2007; Aubry et al. 2008). Diese Einfügung von sechs Nukleotiden könnte anzeigen, wo ein Transposon entfernt wurde (Scott et al. 1996).
Das dritte Merkmal war die Farbe der Samenschale – die auch die Blütenfarbe beeinflusst. Die Blüten sind entweder violett oder weiß. Die weiße Form ist rezessiv (a/a). Sie ergibt sich aus einer Mutation, und zwar einer Transition von Guanin nach Adenin bei einer Spleißstelle* im A-Gen, was zu einem verkürzten Transkriptionsfaktor* führt. Dieser wird verwendet, um die Expression von Genen zu regulieren, die für die Synthese von Anthocyanin (violettem Pigment) erforderlich sind (Ellis et al. 2011). Das vierte, fünfte und sechste Merkmal wurde auf der molekularen Ebene noch nicht untersucht. Das siebte Merkmal war die Stängellänge. Die Pflanzen haben entweder eine normale Höhe oder sind zwergwüchsig. Die Zwergform ist rezessiv (le/le). Sie ergibt sich aus einer Transition von Guanin nach Adenin, die zu einer Substitution der Aminosäure Alanin zu Threonin führt, wodurch ein 3b-Hydroxylase-Enzym (= 3-Oxidase) nicht mehr in der Lage ist, aktives Gibberellinhormon zu bilden, was das Wachstum verhindert (Lester et al. 1997; Martin et al. 1997).
Das sind vier lehrreiche Beispiele. Drei Änderungen ergeben sich aus einem Informationsverlust und unterstreichen damit, wie schädlich Mutationsereignisse sind. Mendels erstes Merkmal ergibt sich jedoch aus einem Transpositionsereignis. In diesem Fall geht die Information nicht verloren, selbst wenn eine Population solcher Pflanzen isoliert und der r/r-homozygote Zustand fixiert war. Denn wenn das Transposon herausspringt, kann die ursprüngliche R-Gen-Funktion wiederhergestellt werden.
Transposons sind allgegenwärtig und auf sie entfällt in den meisten Organismen ein sehr großer Anteil der DNA. Man kann vernünftigerweise davon ausgehen, dass sie bei der phänotypischen Vielfalt eine bedeutsame Rolle spielen.
Die Quellen zu diesem Kastentext finden Sie unter https://mendel-2.pdf
1 Das Problem besteht darin, dass mehrere vorteilhafte Mutationen gleichzeitig erforderlich sind, um eine neue Funktion zu ermöglichen. Wenn sie sukzessiv auftreten, müssten sie aufeinander abgestimmt und jederzeit selektierbar sein. Das ist theoretisch nicht zu erwarten und in der Praxis nie beobachtet worden – im Gegenteil, wie die genannten Beispiele zeigen. Denn mit jeder neuen vorteilhaften Mutation ist die Wahrscheinlichkeit einer weiteren passenden Mutation nicht additiv weniger wahrscheinlich, sondern sogar exponentiell weniger wahrscheinlich und damit sehr bald praktisch unmöglich.
A suggestion as to the nature of “walnut” comb in fowls. Proc. Camb. Phil. Soc. 13, 165–168.
Sonic Hedgehog-signalling patterns the developing chicken comb as revealed by exploration of the Pea-comb mutation. PLoS One 7: e50890.
The genomic substrate for adaptive radiation in African cichlid fish. Nature 513, 375–381.
Wood anatomy of Macaronesian and other Brassicaceae. Aliso 7, 365–384.
Island Biology. Columbia University Press, New York, NY.
On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life. John Murray, London.
The Rose-comb mutation in chickens constitutes a structural rearrangement causing both altered comb morphology and defective sperm motility. PloS Genet 8: e1002775.
Flowering-time genes modulate meristem determinacy and growth form in Arabidopsis thaliana. Nature Genet. 40, 1489–1492.
Versuche über Pflanzenhybriden. Verh. Naturforsch. Vereins Brünn 4, 3–47.
Hybridisierung und Verwandtschaftsgrade innerhalb der Anatidae — eine systematische und evolutionstheoretische Betrachtung. J. Ornithol. 123, 357–380.
Deleterious mutation accumulation and the regeneration of genetic resources. Proc. Natl. Acad. Sci. 95, 394–399.
Germline mutation rates and the long-term phenotypic effects of mutation accumulation in wild-type laboratory mice and mutator mice. Genome Res. 25, 1125–1134.
The industrial melanism mutation in British peppered moths is a transposable element. Nature 534, 102–105.
Parthenocarpic apple fruit production conferred by transposon insertion mutations in a MADS-box transcription factor. Proc. Natl. Acad. Sci. 98, 1306–1311.
Themen | Kurzbeiträge | Streiflichter
Studiengemeinschaft WORT und WISSEN e.V.
Letzte Änderung: 6/21/21
Webmaster