
Themen • Kurzbeiträge • Streiflichter
Gewebereste und Zellbausteine in Dinosaurier-Fossilien
von Harald Binder
Studium Integrale Journal
27. Jahrgang / Heft 1 - Mai 2020
Seite 29 - 36
Zusammenfassung: Organische Gewebereste und sogar Proteinreste wurden in fossilen Dinosaurierknochen nachgewiesen. Diese wurden aus Sedimentschichten geborgen, denen ein Alter von über 60 Millionen radiometrischen1 Jahren zugesprochen wird; das ist völlig unerwartet und steht im Widerspruch zu den Erfahrungen, die mit diesen Stoffen im Labor gemacht werden. Durch beharrliche Arbeit haben Wissenschaftler diese überraschenden Befunde erhärtet. Wie ist die Spannung zwischen bisherigen Erfahrungen und den neuen Ergebnissen zu erklären?
• • • • • • • • • • •
Einführung
Dinosaurier repräsentieren für viele Zeitgenossen eine spannende Epoche der Geschichte der Lebewesen auf der Erde. Diese Geschichte ist uns indirekt durch Hinweise zugänglich, die in den verschiedenen Disziplinen der Geowissenschaften untersucht, beschrieben und interpretiert werden, z. B. Fossilien, geologische Schichten oder Isotopenverteilungen chemischer Elemente. Da eine direkte Beobachtung nicht möglich ist, können wir nicht wissen, wie exakt unsere derzeitige Kenntnis erdgeschichtlicher Abläufe ist. Beispielsweise ist nicht bekannt, wie vollständig die damalige Lebenswelt fossil repräsentiert ist.
Mit einem Stern* versehene Begriffe werden im Glossar erklärt.
Kompakt
Die amerikanische Paläontologin Mary Schweitzer und ihre Mitarbeiter, aber auch andere internationale Arbeitsgruppen, haben seit über 15 Jahren in außerordentlich gut erhaltenen Dinosaurierfossilien flexible und elastische Gewebereste und sogar Bruchstücke von Eiweißen nachgewiesen. Durch unterschiedlichste analytische Nachweismethoden haben sie Messdaten vorgelegt, die es sehr wahrscheinlich machen, dass tatsächlich organische Komponenten aus den Lebewesen, die im Mesozoikum* gelebt haben, erhalten geblieben sind. Damit würden molekulare Informationen zur Verfügung stehen, die uns erlauben, diese Daten aus der Erdgeschichte mit denen aus heutigen Lebewesen zu vergleichen.
Eine große Herausforderung besteht darin, dass bisher noch keine plausible Erklärung für die Erhaltung von empfindlichen Körperbestandteilen von Dinosauriern über die Zeiträume der Erdgeschichte vorliegen. Viele Autoren vermuten noch unbekannte Konservierungsmechanismen, andere schlagen vor, dass Eisen-Ionen, Mineraloberflächen oder Bräunungsreaktionen von Bedeutung gewesen sein könnten. Die Befunde weisen darauf hin, dass wir im Blick auf die Erdgeschichte und die erhaltenen Fossilien grundlegende Aspekte noch wenig verstanden haben.
Vom Organismus zum Fossil – einige grundsätzliche Vorüberlegungen
Unter stabilen ökologischen Bedingungen sollte ein Lebewesen keine körperlichen Spuren hinterlassen.
Mit dem Eintritt des Todes erlischt das typische Verhalten des Lebewesens und seine Körperfunktionen gehen unwiederbringlich verloren. Der Körper des Lebewesens liegt unmittelbar nach Eintritt des Todes zwar in einer Form vor, die von dem kurz zuvor noch lebenden Organismus praktisch nicht zu unterscheiden ist; die organischen Strukturen zerfallen jedoch und lösen sich auf, der Körper geht in die Verwesung über. Dieser natürliche Zerfallsprozess kann beeinflusst werden, in dem man die körperlichen Überreste z. B. einbalsamiert und dadurch Körper mumifiziert. In natürlicher Umgebung kann der Verwesungsprozess unter bestimmten Bedingungen ebenfalls verlangsamt oder blockiert werden, z. B. bei Moor- oder Gletscherleichen, d. h. durch tiefe Temperaturen oder spezielle chemische Randbedingungen (Abwesenheit von Sauerstoff, was bewirkt, dass z. B. viele auf O2 angewiesene Mikroorganismen fehlen, die an der Verwesung beteiligt sind).
Unter natürlichen Bedingungen sollte ein Lebewesen langfristig also keine körperlichen Spuren hinterlassen, vor allem in einem Lebensraum, der ökologisch stabil ist und in dem ein Kadaver den Verwesungsprozessen – einschließlich Aasfressern – zugänglich bleibt.
Anders sieht es aus, wenn irgendeine Konstellation oder eine Störung des Lebensraums die Verwesungsprozesse unterbricht. Die Ursachen und das Ausmaß solcher Störungen während der Erdgeschichte sind uns in der Regel unbekannt. Wir können nur nach geologischen Indizien für mögliche Faktoren suchen, die dazu geführt haben könnten, dass wir Spuren ehemaliger Lebewesen ausgraben können.
Glossar
Femur: Oberschenkelknochen. Histologie: das gr. Wort histos steht für Webstuhl, Kette, Gewebe. Histologie ist die Lehre vom körperlichen Gewebe. Immunochemie: In der Immunochemie werden die biochemischen Prozesse und Strukturen untersucht, die in Organismen zwischen körpereigenen und -fremden Komponenten unterscheiden (Immunreaktion). Kompositmaterial: Vereinigung unterschiedlicher Materialien, wobei die Komponenten ihre jeweilige Mikrostruktur beibehalten und dennoch im Verbund neue Eigenschaften zeigen (z. B. Stahlbeton). Kovalent: Durch gemeinsam genutzte Elektronen (Atombindung) miteinander verknüpfte Atome bzw. Moleküle. marin: Zum Meer gehörig, z. B. marines Lebewesen. Mesozoikum: Sogenanntes „Erdmittelalter“, geologische Periode mit den Formationen Trias, Jura und Kreide. Die hier diskutierten Fossilfunde stammen aus der Oberkreide. MrJ: Millionen radiometrische Jahre (Altersbestimmung mithilfe der Radiometrie).
Sehr gut erhaltene Fossilien mit mikroskopischen Strukturen
Wenn wir Fossilien von Dinosauriern, diesen besonderen Echsen aus dem Mesozoikum*, in sehr guter Erhaltung finden, sogar mit mikroskopisch auflösbaren Strukturen, dann ist das ein seltener Glücksfall. Über die Bedingungen, die zu einer solch exzellenten Erhaltung führen, wissen wir bisher sehr wenig. 1966 veröffentlichten Pawlicki und Mitarbeiter mikroskopische Befunde von Kollagenstrukturen und Knochenzellen in fossilen Dinosaurierknochen (s. auch Schweitzer & Horner 1999 und dort zitierte Arbeiten). In den Veröffentlichungen zu diesen außerordentlich gut erhaltenen Fossilien beschreiben und dokumentieren die Autoren auch Gewebe- und Gefäßstrukturen aus dem Inneren, dem schwammartigen, porösen Bereich der massiven Knochen. Knochen und Zähne sind Körperbestandteile, die im Vergleich zum Gewebe der Weichteile gute Voraussetzungen bieten, fossil erhalten zu werden. Sie stellen bezüglich ihrer materiellen Zusammensetzung sogenannte Kompositmaterialien* dar. Sie bestehen aus anorganischen, mineralischen Komponenten, die eine Art Gerüst oder eine Matrix darstellen, in der organische Komponenten enthalten sind. In Knochen sind auch Zellen eingelagert (Osteozyten und -klasten), die den Knochen auf- und abbauen.
Die eben genannten ersten Veröffentlichungen fanden keine besondere Beachtung. Im Vergleich dazu glich die Veröffentlichung von Schweitzer et al. (2005) einem Paukenschlag. Diese Arbeit über kleine, flexible Gewebereste (Abb. 1), die sie und ihr Mitarbeiterteam aus dem fossilen Femur (Oberschenkelknochen) eines Tyrannosaurus rex gewinnen konnten, erregten großes Aufsehen. Bevor wir darauf zu sprechen kommen, sollen einige Angaben zur zeitlichen Einordnung der Funde gemacht werden.
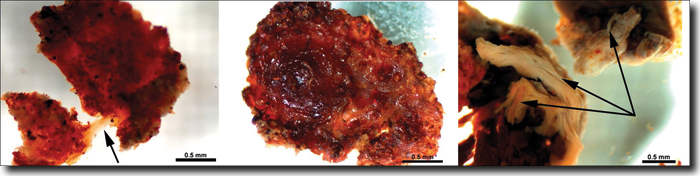
Abb. 1: Flexible und elastische Gewebepartikel aus dem Femur des Tyrannosaurus rex, nachdem Proben aus dem Bereich des Knochenmarks demineralisiert worden waren. (Schweitzer et al. 2005)
Altersangaben für die mesozoischen Fossilien
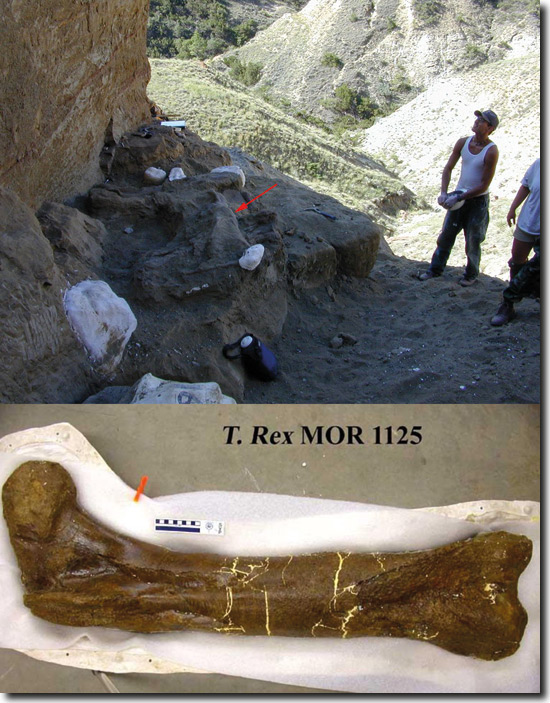
Abb. 2: Fundsituation bei der Bergung des fossilen T. rex-Femur (weißer Pfeil) und nach erfolgter Präparation. (Foto Fundstelle: J. R. Horner; Museum of the Rockies, Montana, USA; mit freundlicher Genehmigung; Foto Femur: Mary H. Schweitzer, mit freundlicher Genehmigung)
Für die beiden Fossilien, die in diesem Artikel hauptsächlich zur Sprache kommen, sind in der Literatur folgende Alter angegeben:
Tyrannosaurus rex: Dieses Fossil wurde aus dem unteren Bereich der Hell Creek Formation in Montana, USA, geborgen, 8 m über dem Fox Hill-Sandstein in der geologischen Formation der Kreide (oberste Formation des Mesozoikums*). Die Fundlokalität ist geowissenschaftlich charakterisiert und datiert; das Alter wird mit ca. 68 MrJ* angegeben (Abb. 2).
Brachylophosaurus canadensis: Der für die Untersuchungen genutzte fossile Femur* dieses Entenschnabelsauriers wurde 2007 unter einer 7 m mächtigen Sandstein-Formation (Judith River-Formation) geborgen. Von einer 10–12 cm dicken Sandsteinmatrix umgeben, wurde das Fossil erst im Labor freipräpariert und unmittelbar danach wurden die Proben genommen. Als radiometrisches Alter geben die Autoren ca. 80 MrJ an.
Es liegen aber auch 14C (Radiokarbon)-Datierungen vor. Von einer Radiokarbon-Datierung von Fossilien aus mesozoischen Sedimentschichten würde man eigentlich ein quasi-unendliches 14C-Alter erwarten, d. h. keine zuverlässig nachweisbaren Mengen an 14C. Weil vom Kohlenstoff-Isotop mit der Masse 14 u nach 5730 +/- 40 Jahren die Hälfte der Atome radioaktiv zerfallen ist (Halbwertszeit), kann man bei einem Probenalter von über 60.000 Jahren wegen der äußerst geringen Menge verbliebenen 14C keine vertrauenswürdigen Daten mehr erwarten. Dennoch haben Miller et al. (2019) Daten aus 14C-Datierungsexperimenten von Saurierfossilien vorgelegt. Die Datierungen ergeben 14C-Alter von 22.000 bis 41.000 radiometrischen Jahren. Diese Befunde werden in aktuellen Fachpublikationen allerdings nicht erwähnt und nicht diskutiert.
Erste überraschende und aufsehenerregende Befunde
Aus Fragmenten, die bei der Bergung eines sehr gut erhaltenen fossilen Femurs eines T. rex angefallen waren, hatte Mary Schweitzer Proben präpariert, um diese mikroskopisch untersuchen zu können. Der Fokus lag dabei vor allem auf der Suche nach Hinweisen auf die Gewebestruktur in den fossilen Knochen. Erste Resultate dieser histologischen* Studien lieferten Hinweise auf rote runde Körperchen innerhalb von Gefäßstrukturen (verzweigte Kanalsysteme; Abb. 3). Dabei entsprachen die Größenverhältnisse denjenigen von roten Blutkörperchen (Erythrozyten). So etwas war zuvor noch nicht beschrieben worden, und das Vorliegen solcher Strukturen entsprach in keiner Weise den Erwartungen. Solche biologischen Systeme können nach bisher etablierten wissenschaftlichen Vorstellungen bei Fossilien aus dem Mesozoikum nicht erhalten bleiben.
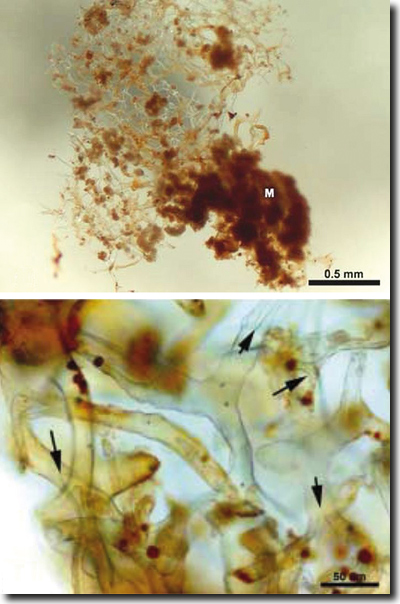
Abb. 3: Nach vollständiger Demineralisierung von Proben aus der Rinde des Femurs von T. rex werden flexible und transparente Gefäßsysteme erkennbar. Diese ähneln weder Pilzhyphen noch Pflanzenteilen. In der Vergrößerung sind in den Hohlräumen der Gefäße runde, rote Körperchen zu erkennen. (Aus Schweitzer et al. 2005)
Es folgten viele Versuche, die gefundenen mikroskopischen Strukturen in dem fossilen T. rex-Femur-Fragment als Artefakte zu erweisen; also als irgendwelche z. B. durch Fossilisationsprozesse erzeugte Formen, die Gefäße und Erythrozyten nur vortäuschen. Alle Bemühungen, die Interpretation der fossilen mikroskopischen Strukturen als Blutgefäße und Erythrozyten zu widerlegen, schlugen fehl und erhärteten im Gegenteil den ursprünglichen Eindruck.
Schließlich wurden pulverisierte Proben des fossilen Materials in ein Lösungsmittel (einen Komplexbildner) gegeben, das die mineralischen Komponenten der Probe auflöst. Im Rückstand der demineralisierten Probe fanden sich Gewebereste, die Größen von bis zu wenigen Millimetern aufwiesen. Diese Gewebereste waren flexibel und elastisch. Außerdem konnten verzweigte Gefäßfragmente mit erythrozytenartigen Körperchen isoliert und dokumentiert werden. Vergleichende Untersuchungen an Knochen von Küken und von Straußen lieferten vergleichbare Daten und unterstützten damit die Interpretation der Befunde als organische Komponenten aus den fossilen T. rex-Knochen. Schweitzer et al. (2005) dokumentieren in ihrer Veröffentlichung auch parallel orientierte Fasern, die zellähnliche Strukturen umgeben. Durch Vergleich mit Präparaten aus Knochen eines rezenten Vogels Strauß deuten die Forscher an, dass es sich dabei um Kollagenfasern und Osteozyten (Knochenzellen) handeln könnte. Abschließend wird in der Publikation erwähnt, dass die Autoren in Extrakten aus den Fossilien Proteine identifiziert haben, die Hinweise auf immunochemische* Reaktionen zeigen.
In dieser aufsehenerregenden Arbeit legen die Autoren somit eine Reihe von Belegen für die völlig unerwartete Erhaltung organischer Bestandteile in fossilisierten Dinosaurierknochen vor. Sie sind in ihren Schlussfolgerungen aber auffallend zurückhaltend, wohl in dem Wissen um die etablierte Vorstellung, dass organische Reste aus der Zeit des Mesozoikums nicht bis in die Gegenwart erhalten werden können.
Die etablierten wissenschaftlichen Vorstellungen sperren sich gegen die Akzeptanz von organischen Resten in Fossilien aus dem Mesozoikum.
Zwei Jahre später konnte die Arbeitsgruppe um Mary Schweitzer weitere Befunde vorlegen, die ihre bisherige Interpretation der Daten unterstützen (Schweitzer et al. 2007). An denselben Fossilien, die bereits für die Arbeiten zur Veröffentlichung von 2005 verwendet wurden, wurden Präparationen mit rasterkraftmikroskopischen Methoden (AFM) untersucht. Damit konnte gezeigt werden, dass die einzelnen Fasern selbst eine sich wiederholende Unterstruktur enthalten, deren Periodizität bei 70 nm lag. Dieses Merkmal passt zum molekularen Aufbau von Kollagenfasern. Präparate aus modernen Knochen eines Emus wiesen in vergleichenden Studien dieselben Merkmale auf. Immunologische Untersuchungen erhärteten die Vermutung, dass Kollagen I in den mesozoischen Dinosaurierfossilien vorliegt. Außerdem wurden typische Komponenten, die am Aufbau von Kollagen I beteiligt sind (wie z. B. die Aminosäuren Glycin und Prolin), mittels Massenspektrometrie nachgewiesen.
Das Bild kann online nicht zur Verfügung gestellt werden.
Abb. 4: Massenspektrometrischer Nachweis von Kollagenfragmenten aus Fossilien von T. rex (68 MrJ) und Mastodon (0,5 MrJ), einer ausgestorbenen Elefantenart. (© Zina Deretsky, National Science Foundation)
In einer Publikation, die in derselben Ausgabe von Science unmittelbar im Anschluss an die Arbeit von Schweitzer et al. (2007) erschien, legt dieselbe Arbeitsgruppe weitere ausführliche Daten von vergleichenden massenspektrometrischen Untersuchungen von Proteinsequenzen aus Fossilien von T. rex und einem Mastodon (Mammut americanum) aus pleistozänen Ablagerungen vor (Asara et al. 2007; Abb. 4). Darin werden Fragmente der Kollagen-Eiweißkette durch Vergleich mit entsprechenden synthetischen Peptiden nachgewiesen. Die beiden zuletzt genannten Publikationen stießen auf großes Interesse, wurden aber auch sehr kritisch und kontrovers diskutiert (s. Binder 2009).
In der Kritik wird vor allem auf den Widerspruch zwischen den neuen Befunden und den etablierten Kenntnissen über die Geschwindigkeit von Zerfallsprozessen und auf die Gefahr von Kontamination verwiesen. Damit wird die Vermutung geäußert, dass die dokumentierten Proteine nicht aus den Dinosaurierknochen stammen, sondern aus der Umgebung der Fossilien oder im Verlauf ihrer Bergung und Präparation oder bei der Analyse aus anderen Quellen hinzugekommen seien. Die analytisch dokumentierten Peptide wären damit Verunreinigungen.
Als weitere Quelle der Proteine werden auch Mikroorganismen angeführt, die als Biofilm bei Fossilisationsprozessen eine Rolle spielen können, im Sediment aber auch während der gesamten Handhabung der Fossilien (Bergung, Präparation usw.) allgegenwärtig sind. Auch im Blick auf einige Ungereimtheiten bei den massenspektrometrischen Untersuchungen und der damit im Zusammenhang stehenden Nutzung von Datenbanken und bioinformatischen Methoden wurde Kritik geübt.
Die Autoren um Mary Schweitzer sind auf die geäußerte Kritik detailliert eingegangen, haben ihre Argumente in den Erwiderungen geschärft und argumentieren bis heute sehr nüchtern und eher zurückhaltend; die Kritik an Methoden und experimentellen Details haben sie in weiteren Arbeiten berücksichtigt und aufgenommen. An ihren Einschätzungen hat sich dadurch nichts Wesentliches geändert.
Das Bild kann online nicht zur Verfügung gestellt werden.
Abb. 5: Femur des Entenschnabelsauriers (Brachylophosaurus) mitsamt der Hülle aus umgebendem Sediment und Gipsbinden, die im Gelände angebracht wurden. Zu sehen ist der Bereich, an dem die Proben für die chemischen Analysen genommen wurden. (© Mary H. Schweitzer)
2009 veröffentlichten Schweitzer et al. die Ergebnisse von Untersuchungen an Fossilien eines Entenschnabelsauriers (Hadrosaurier) Brachylophosaurus canadensis. Diese waren 2007 unter großen Vorsichtsmaßnahmen mit dem umgebenden Sediment (Sandsteinmatrix) geborgen und unter sterilen Bedingungen im Labor präpariert worden (Abb. 5). Mit den in dieser Arbeit dokumentierten Daten konnten die Autoren zeigen, dass die an den T. rex gewonnenen Resultate kein Einzelfall waren. Auch an diesen außerordentlich gut erhaltenen Fossilien konnten nach Demineralisierung mikroskopisch faserige Strukturen in der Knochenmatrix nachgewiesen werden. Verzweigte Gefäße mit roten runden Körperchen darin wurden ebenso gefunden wie auch Knochenzellen mit ihren charakteristischen Ausstülpungen, den Filipodia. In chemischen Extrakten konnten Kollagen I und weitere Proteine (Laminin und Elastin, die ebenfalls Bestandteile von Blutgefäßen sind) immunologisch nachgewiesen werden. Der immunohistochemische Nachweis gelang auch direkt an den demineralisierten Präparaten durch sogenannte „in situ“-Analysen. Massenspektrometrisch konnten an den Fossilien von B. canadensis acht Peptidsequenzen dokumentiert werden, die bis zu max. 33 Aminosäuren enthielten.
2014 untersuchten Cleland et al. gezielt Proteine aus flexiblen Gefäßen von mesozoischen Fossilien. Dazu hatten sie aus den Fossilien von B. canadensis die zuvor beschriebenen Gefäße isoliert. Daran erhielten sie positive immunochemische Hinweise auf die Strukturproteine Actin, Tubulin, Myosin und Tropomyosin. Sie konnten deren Vorkommen aufgrund von Proteinfragmenten auch massenspektrometrisch aufzeigen. Die erhaltenen Peptidsequenzen passen zu jenen, die aus Knochen vom heute lebenden Huhn und Vogel Strauß gewonnen wurden, und nicht zu Proteinen aus Bakterien, Biofilmen und Pilzen. Außerdem stammten die massenspektrometrisch bestimmten Proteinfragmente genau aus den Bereichen der Fossilien, in denen mittels Antikörper Proteinstrukturen lokalisiert worden waren.
2017 veröffentlichte ein Autorenteam um Mary Schweitzer eine Wiederholung der Analysen an den Fossilien des Hadrosauriers von 2009. Dabei wurden die in der Zwischenzeit gewonnenen Erfahrungen und weiterentwickelte Methoden in der Probenherstellung und -präparation angewendet (Schroeter et al. 2017). In dieser Studie konnten wieder acht Kollagen I-Fragmente erfasst werden, davon waren zwei identisch mit den 2009 nachgewiesenen. Sechs Peptidsequenzen waren neu. Mit diesen Befunden liefern die Autoren erneut Argumente dafür, dass in Fossilien aus der Kreide Proteinfragmente erhalten sind, die heute analytisch nachgewiesen werden können.
Nachweis von Proteinfragmenten in gut erhaltenen Dinosaurierfossilien
Bei den Bemühungen um einen Nachweis, dass die gefundenen Proteinfragmente ursprünglich zum lebenden Organismus gehörten, also endogen sind und an dessen Aufbau beteiligt waren, wurden bisher vor allem die beiden Strukturproteine Kollagen und Keratin sehr intensiv untersucht.
Kollagen in Fossilien
Unter Kollagen werden verschiedene Proteine zusammengefasst, die im Körper von Tier und Mensch für Festigkeit sorgen und die man daher als Struktur- oder Skleroproteine bezeichnet (skleros (gr.) = trocken, hart, rauh). Kollagen, ein faserbildendes Protein, findet sich in großen Mengen (ca. 30 % des Proteinanteils im menschlichen Körper) in Knochen, Zähnen, Sehnen, Bändern und der Haut (also vor allem in dem, was man gemeinhin als Bindegewebe bezeichnet, einschließlich der beim Menschen in jüngerer Zeit so viel beachteten Faszien).
Kollagen wird in unterschiedliche Typen eingeteilt, Kollagen I ist der häufigste Typ. Die einzelnen Proteinketten können aus einigen Hundert bis wenigen Tausend Aminosäuren bestehen. In großen Bereichen (sog. Domänen) weist die Abfolge der Aminosäuren ein typisches, sich wiederholendes Triplett der Form G-X-Y auf. Dabei steht G für die kleinste Aminosäure Glyzin, X für die Aminosäure Prolin; das Y steht für ein zweites Prolin, das mit einer OH-Gruppe versehen ist und Hydroxyprolin genannt wird. Aus drei Proteinketten wird eine rechtsgängige Helix, eine Tripelhelix, gebildet, die man als Tropokollagen bezeichnet. Die Tripelhelix hat einen Durchmesser von ca. 1,5 nm (1 nm ist 1 Millionstel Millimeter). Die Kollagen-Tripelhelices können zu sogenannten Mikrofibrillen zusammengelagert werden, in denen die einzelnen Tropokollagen-Bausteine regelmäßig, etwa um ein Fünftel ihrer Länge (67 nm) gegeneinander verschoben, aneinander gelagert sind. Die Mikrofibrillen haben einen Durchmesser von 20-40 nm. Die Mikrofibrillen können sich ihrerseits zu noch größeren Aggregaten, den Fibrillen (300 bis 500 nm Durchmesser), zusammenlagern und diese erneut zu Fasern (4-12 µm Durchmesser).
Die durch die gestaffelte Zusammenlagerung der Tripelhelices in den Mikrofibrillen bewirkten Dichteunterschiede können als typisches Muster der Fibrillen elektronenmikroskopisch dargestellt werden. Auf diese Weise kann Kollagen mikroskopisch nachgewiesen werden.
Die besonders dichte Packung der Kollagenproteine und die kovalente* Verknüpfung der Tropokollagen-Tripelhelices in den Mikrofibrillen sind die Ursache dafür, dass Kollagen als Strukturprotein im Vergleich zu anderen Proteinen robust ist. Kollagen ist also gegenüber Abbauprozessen (Hydrolyse) oder auch enzymatischem Abbau widerstandsfähiger als z. B. viele Enzyme, deren Aktivität aufgrund struktureller Veränderungen in Pufferlösungen und durch Hydrolyse z. T. sehr rasch abfallen kann. In vielen Tieren, auch in Menschen und Mikroorganismen, sind Enzyme – Proteasen – bekannt, die die Bindung zwischen Prolin und anderen Aminosäuren spalten und als Kollagenasen bezeichnet werden.
Weitere Befunde
Lindgren et al. (2011) untersuchten einen sehr gut erhaltenen fossilen Oberarmknochen (Humerus) eines Masosauriers (ausgestorbenes, im Meer lebendes Reptils, auch als Maasechse bezeichnet) aus einem marinen* Sediment, das der Kreide zugeordnet wird (70 MrJ). Durch Kombination verschiedener Techniken – optische Mikroskopie, Elektronenmikroskopie, Immunochemie, Massenspektrometrie und IR-Spektrometrie – konnten sie überzeugende Argumente liefern, dass organische Moleküle, einschließlich Kollagenfragmente, in den faserigen Geweberesten des fossilen Humerus vorliegen. In dieser und auch den folgenden Studien wurden die organischen Komponenten nicht extrahiert, sondern direkt in den mikroskopisch charakterisierten erhaltenen Geweberesten nachgewiesen.
Eine chinesisch-taiwanesische Arbeitsgruppe hat aus unteren jurassischen Sedimentschichten (Dawa, Provinz Yunnan, China) geborgene Fossilien eines Echsenbeckensauriers (Sauropodomorpha), Lufengosaurus, auf Hinweise nach Kollagen untersucht (Lee et al. 2017). Für das Alter geben die Autoren 190 bis 197 MrJ an. In einer fossilisierten Rippe dokumentieren sie mikroskopisch Blutgefäße, aus denen sie transparentes Material auswaschen konnten. Darin wiesen sie mit einer speziellen Methode der Infrarot-Spektroskopie, in der sie Synchrotron-Strahlung nutzen, Proteinfragmente nach. In diesen Gefäßen fanden sie auch Kügelchen aus Hämatit (Durchmesser: 6–8 µm), ein Eisenoxid-Mineral (α-Fe2O3). Die Autoren äußern die Vermutung, dass das Eisenmineral für die Konservierung eine wichtige Rolle spielen könnte. 2013 legte dasselbe Autorenteam eine vergleichbare Untersuchung an einem in China gefundenen fossilen Dinosaurierembryo vor (Unterjura, Süden Chinas; 190–197 MrJ; Reisz et al. 2013). Auch in dieser Studie konnten sie organische Proteinfragmente nachweisen.
Surmik et al. (2016) publizierten eine Untersuchung an Fossilien von im Meer und an Land lebenden Sauriern, die aus Sedimentgestein der frühen und mittleren Trias (247,2 MrJ) in Polen geborgen worden waren. Auch sie dokumentieren Blutgefäße aus fossilisiertem Knochen. Darin weisen sie durch eine Kombination von mikroskopischen und spektroskopischen Methoden Aminosäuren nach, die für das Vorhandensein von Kollagenfragmenten sprechen. Die Autoren nehmen in Anspruch, damit den bisher ältesten Nachweis von komplexen organischen Molekülen aus Fossilien von Wirbeltieren aus marinen Lebensräumen beschrieben zu haben.
Rezeption der Funde von Kollagen in Dinosauriern aus dem Mesozoikum
Inzwischen liegen zahlreiche Veröffentlichungen von Studien vor, in denen Proteinstrukturen in Fossilien nachgewiesen werden, was bei den jeweils zugeordneten Altern aufgrund von Laborerfahrungen nicht zu erwarten ist. Sie erhärten den Eindruck, dass die Interpretation der Proteinreste als vom ursprünglichen Lebewesen stammend plausibel ist.
Dennoch gibt es nach wie vor kritische Stimmen, die als Ursache für die Protein-Nachweise Kontaminationen vermuten. Die Gefahr von Verunreinigungen ist während der Bergung, der Präparation und auch während der Handhabung im Labor gegeben. Auch werden von manchen Autoren nach wie vor Mikroorganismen bzw. aus ihnen gebildete Biofilme als eigentliche Quelle der Proteine angesehen (Saitta et al. 2019, Buckley et al. 2017), eben weil die Erhaltung von Proteinstrukturen, aufgrund von Erfahrungen im Umgang mit Proteinen im Labor, über so lange Zeiten nicht zu erwarten sei.
Um zukünftige Untersuchungen von Proteinen oder überhaupt organische Biomoleküle aus Fossilien besser abzusichern, haben verschiedene Arbeitsgruppen Beiträge geleistet und Vorsichtsmaßnahmen zusammengestellt und veröffentlicht (Hendy et al. 2018, Schweitzer et al. 2019). Damit sollen die Befunde zukünftiger Studien an Vertrauenswürdigkeit gewinnen.
Auf der Suche nach plausiblen Erklärungen
Mary Schweitzer und andere Kollegen sind intensiv auf der Suche nach möglichen Erklärungen für das Vorkommen von Gewebestrukturen und Zellbestandteilen in mesozoischen Fossilien. Man vermutet diese Erklärungen in der kompakten Struktur der Kollagenfibrillen oder auch in günstigen geochemischen Randbedingungen. Jüngst wurden auch Bräunungsreaktionen (Maillard-Reaktionen) angeführt. Die bisher geäußerten Vorschläge ändern nichts daran, dass die Erhaltung von organischen Makromolekülen über einen Zeitraum von über 60 Millionen Jahren bisher nicht verstanden ist.
Diskussion
Schweitzer et al. (2009) schreiben, nachdem ihre erste Veröffentlichung über flexible und elastische Gewebereste in T. rex-Fossilien aus der Kreide (Schweitzer et al. 2005) kontrovers diskutiert worden war: „Die einfachste (sparsamste) und bisher nicht widerlegte Erklärung ist, dass in manchen Fossilien von Dinosauriern aus der Kreide ursprüngliche Moleküle [eigentlich: Fragmente davon; H. B.] erhalten geblieben sind. Die Chemie, die diese Erhaltung bewirkt, ist noch unbekannt.“2 Die Hypothese, dass Proteinstrukturen in Fossilien aus der Kreide und aus noch älteren Sedimentgesteinen heute noch analytisch nachgewiesen werden können, ist durch mehrere Befunde verschiedener Arbeitsgruppen erhärtet worden. Allerdings gilt auch noch der zweite Satz aus dem Zitat, dass bis heute noch kein plausibler Mechanismus aufgezeigt worden ist, der diese Befunde verständlich machen würde.
Die aus Labordaten abgeschätzte Lebensdauer von Kollagen (Buckley et al. 2008) beläuft sich maximal auf wenige 100.000 Jahre unter günstigen Bedingungen (wie niedere Temperatur, wenig Feuchtigkeit und weitgehende Abwesenheit von Mikroorganismen).
Eingangs der Veröffentlichung von Schweitzer et al. (2014) bemerken die Autoren: „Die Langlebigkeit von ursprünglichem Gewebe in fossilen Knochen aus dem Mesozoikum wird durch aktuelle chemische Modelle für deren Abbau nicht erklärt.“3 Viele Autoren postulieren mit Mary Schweitzer einen derzeit noch unbekannten Mechanismus zu Konservierung von Proteinfragmenten in mesozoischen Fossilien. Bis ein solcher Mechanismus aufgezeigt und gut begründet ist, stellt das eine Spekulation dar, die zukünftig zu prüfbaren Hypothesen zugespitzt werden sollte. Selbst wenn im Labor Konservierungsmechanismen aufgezeigt werden könnten, wissen wir dadurch nicht, was sich bei den Fossilisationsprozessen im Verlauf der Erdgeschichte wirklich abgespielt hat, denn die Vergangenheit ist unserer Empirie nicht direkt zugänglich.
Mechanistisch ist die Erhaltung von organischen Makromolekülen über einen Zeitraum von über
60 Millionen Jahren nicht verstanden.
Grundsätzlich ist anzumerken, dass das Erklärungsdefizit für die Erhaltung von Proteinfragmenten in Fossilien aus dem Mesozoikum erheblich verringert würde, wenn die dem Mesozoikum und dem Känozoikum zugeschriebenen Alter komprimiert würden. Denkansätzen dieser Art stehen etablierte Vorstellungen zur Erdgeschichte entgegen. Aber die noch junge Geschichte der Erforschung von Proteinfragmenten in alten Fossilien lehrt: Es gibt Beobachtungen, die bei sorgfältiger und intensiver Prüfung etablierte Vorstellungen überwinden können. Dies gilt in dem hier verhandelten Fall auch dann, wenn noch großer Erklärungsbedarf besteht. Mary Schweitzer hat mit ihren Kollegen eindrucksvoll gezeigt, dass durch anhaltende und sorgfältige Forschung etablierte Betrachtungsweisen herausgefordert und überwunden werden können.
Insgesamt aber zeigen die hier vorgestellten Forschungen, dass wir im Blick auf ein Verständnis der Erdgeschichte immer wieder mit Überraschungen rechnen müssen und offen bleiben müssen für neue Befunde, ihre Erklärung und ihr Verständnis.
Mendel zeigte, dass die Genetik quantisiert ist, d. h. in „Pakete“ gegliedert, und wir wissen heute, dass die individuellen Informationspakete durch Gene definiert sind. Dominante und rezessive Merkmale werden dann einfach als das Vorhandensein und Fehlen von funktionalen Genen verstanden. Wird ein defektes Gen von nur einem Elternteil vererbt, kann es durch das Gen des anderen Elternteils kompensiert werden. Das läuft darauf hinaus, dass eine Merkmalsausprägung, die mit einem bestimmten Gen verbunden ist, dominant ist. Ein rezessives Merkmal wird erst dann ans Licht gebracht, wenn ein Individuum von beiden Elternteilen defekte Kopien erhält.
Diese neuen Erkenntnisse haben schwerwiegende Auswirkungen auf unser Verständnis von Biologie. Wenn Gene verloren gehen müssen, damit rezessive Merkmale ausgeprägt werden können, wie sind dann diese Gene ursprünglich entstanden? Offensichtlich gibt es im Genom viel mehr Gene, als sich durch die bekannten Evolutionsmechanismen erklären lassen. Die Gesetze von Mendel zeigen sich bei Genen, die für das Überleben des Organismus nicht unbedingt notwendig sind. Sie dürfen sogar defekt oder auch gar nicht vorhanden sein, ohne dass dies zu schweren Schäden führt. Verlustmutationen sind der Grund, warum Mendel kurze und lange Pflanzen, weiße und rosa Blüten, gelbe und grüne Samen fand. Verlust von Genen bzw. Allelen (Homozygotisierung) ist ein eleganter Mechanismus, um Variationen, Anpassungen und neue Arten zu erzeugen. Da es sich aber um einen Zerfallsmechanismus handelt, sprechen die Zerfallstendenzen des Erbguts (genetische Entropie) dagegen, dass auf diesem Wege neue genetische Information hervorgebracht werden kann.
Hinweise: Eine ausführlichere Version dieses Artikels ist als Internetartikel veröffentlicht unter: https://www.wort-und-wissen.org/wp-content/uploads/b-20-2_dinosaurier.pdf
1 In den Fachpublikationen wird das auf Messung der Verhältnisse von Radioisotopen basierende Alter als kalendarisches Alter dargestellt. Dabei werden die so bestimmten radiometrischen Alter also mit Kalenderjahren gleichgesetzt. Im Text wird durch den Zusatz „radiometrisch“ auf diesen Sachverhalt hingewiesen.
2 „The most parsimonious explanation, thus far unfalsified, is that original molecules persist in some Cretaceous dinosaur fossils. Still unknown is the chemistry behind such preservation.“
3 „The persistence of original soft tissue in Mesozoic fossil bone is not explained by current chemical degradation models.“
The half-life of DNA in bone: measuring decay kinetics in 158 dated fossils. Proc. R. Soc. B doi: 10.1098/rspb.2012.1745
Protein sequences from Mastodon and Tyrannosaurus rex revealed by mass spectrometry. Science 316, 280–285.
Dinosaur paleohistology: review, trends and new avenues of investigation. PeerJ, doi:10.7717/peerj.7764.
Fibres and cellular structures preserved in 75-million-year-old dinosaur specimens. Nat. Comm. 6:7325.
Neue Befunde zu alten Proteinen aus Dinosaurierfossilien. Stud. Int. J. 16, 104–107.
Weighing the mass spectrometric evidence for the authentic Tyrannosaurus rex collagen. Science 319, 33.
A fossil protein chimera; difficulties in discriminating dinosaur peptide sequences from modern cross-contamination. Proc. R. Soc. B 284:20170544; doi: 10.1098/rspb.2017.0544.
Mass spectrometry and antibody-based characterization of blood vessels from Brachylophosaurus canadensis. J. Proteome Res. Doi: 10.1021/acs.jproteome.5b00675.
A comparison of common mass spectrometry approaches for paleoproteomics. J. Prot. Res. 17, 936–945.
Protein sequences bound to mineral surfaces persist into deep time. eLife 2016;5:e17092; doi: 10.7554/eLife.17092.
A guide to ancient protein studies. Nature Ecol. Evol. 2, 791–799.
Evidence of preserved collagen in an early Jurassic sauropodomorph dinosaur revealed by synchroton FTIR microspectroscopy. Nat. Commun. 8:14220; doi: 10.1038/ncomms14220.
Ausbruch des Laacher-See-Vulkans. Stud. Int. J. 26, 19–26.
Microspectroscopic evidence of Cretaceous bone Proteins. PlosOne 6: e19445; doi:10.1371/journal.pone.0019445.
The search for solutions to mysterious anomalies in the geologic column. Geol. Earth Mar Sci. 1, 1–15.
Cells, collagen fibrils and vessels in dinosaur bone. Nature 211, 655–657.
Embryology of early Jurassic dinosaur from China with evidence of preserved organic remains. Nature 496, 210–214.
Not so dry bones: An interview with Mary Schweitzer on July 21, 2014; https://biologos.org/articles/not-so-dry-bones-an-interview-with-mary-schweitzer (abgerufen am 18. 3. 2020).
Dinosaurpeptides suggest mechanisms of protein survival. PLOSone 6: e20381; doi: 10.1371/journal.pone.0020381.
Cretaceous dinosaur bone contains recent organic material and provides an environment conductive to microbial communities. eLife 8: e46205; doi: 10.7554/eLife.46205.
Expansion for the Brachylophosaurus canadensis Collagen I sequence and additional evidence of the preservation of Cretaceous protein. J. Proteome Res.; doi: 10.1021/acs.jproteome.6b00873.
Heme compounds in dinosaur trabecular bone. Proc. Natl. Acad. Sci. 94, 6291–6296.
Intravascular microstructures in trabecular bone tissue of Tyrannosaurus rex. Ann. Paléontol. 85, 179–192.
Identification of immunoreactive material in mammoth fossils. J. Mol. Evol. 55, 696–705.
Soft-tissue vessels and cellular preservation in Tyrannosaurus rex. Science 307, 1952–1955.
Blutspuren aus der Kreidezeit. Spektrum der Wissenschaft; https://www.spektrum.de/news/blutspuren-aus-der-kreidezeit/1072452.
Analysis of soft tissue from Tyrannosaurus rex suggest the presence of protein. Science 316, 277–280.
Biomolecular characterization and protein sequences of the campanian Hadrosaur B. Canadensis. Science 324, 626–631.
A role of iron and oxygen chemistry in preserving soft tissues, cells and molecules from deep time. Proc. R. Soc. B 281:20132741; doi: 10.1098/rspb.2013.2741.
Paleoproteomics of mesozoic dinosaurs and other Mesozoic fossils. Proteomics doi: 10.1002/pmic.201800251.
Keeping the faith. Science 357, 1088–1091.
Cretaceous dinosaur bone contains recent organic material and provides an environment conductive to microbial communities. eLife 8; doi: 10.7554/eLife.46205.
Spectroscopic studies on organic matter from Triassic reptile bones, Upper Silesia, Poland. PLOSone 11; e0151143; doi:10.1371/journal.pone.0151143.
Themen | Kurzbeiträge | Streiflichter
Studiengemeinschaft WORT und WISSEN e.V.
Letzte Änderung: 6/25/21
Webmaster