
Themen • Kurzbeiträge • Streiflichter
Mendel‘sche Artbildung und die Entstehung der Arten
1. Woher kommt die Artenvielfalt?
2. Latente Information und präexistente genetische Programme
3. Fortpflanzungsisolation und adaptive Radiationen
von Nigel Crompton
Studium Integrale Journal
26. Jahrgang / Heft 2 - Oktober 2019
Seite 86 - 92
Zusammenfassung: Wie kann Vielfalt innerhalb von genetischen Familien bzw. Grundtypen entstehen? Der durch seine Vererbungsgesetze bekannte Gregor Mendel entwickelte zu dieser Frage eine Erklärung, die heute nahezu vergessen ist. Aber gerade neuere Befunde über rasche Artbildung sind starke Indizien dafür, dass nicht Darwin, sondern Mendel Recht hatte: Artbildung „lebt“ von präexistenten genetischen Programmen.
• • • • • •
Einleitung
Die Frage, wie Artbildung erfolgt, also die Aufspaltung von einer Art in zwei oder mehr Tochterarten, gehört zu den zentralen Fragen der Biologie. Charles Darwins epochemachendes Werk „On the Origin of Species“ trägt diese Frage implizit im Titel. Im Gefolge Darwins sollen Arten sich dadurch auseinanderentwickeln und aufspalten, dass sich nach und nach Mutationen* ansammeln, Populationen geographisch getrennt werden und in den getrennten Populationen unterschiedliche Selektion wirkt. Im Laufe der Zeit sollen schließlich Fortpflanzungsbarrieren entstehen (genetische Isolation). Auf dieser Basis werden große Zeiträume für die Bildung neuer Arten veranschlagt. Dieses Verständnis ist heute Mainstream in der Evolutionsbiologie.
Mit einem Stern* versehene Begriffe werden im Glossar erklärt.
Abb. 1: Gregor Johann Mendel, der Vater der modernen Genetik. Geboren am 20. Juli 1822 in Heinzendorf, Schlesien (heute Hyncice, Tschechien). Er nahm den Namen Gregor an, nachdem er Augustinermönch in der Abtei St. Thomas, Brünn, Österreich (heute Brno, Tschechien), geworden war. 1868 wurde er dort Abt. Er führte seine Experimente mit Pflanzenhybriden zwischen 1854 und 1865 durch und veröffentlichte 1866 seine berühmte Arbeit. Er starb am 6. Januar 1884 in Brünn im Alter von 61 Jahren. (Mit freundlicher Genehmigung der Abtei St. Thomas, Brno)
Ein Zeitgenosse Darwins, der Augustinermönch Gregor Mendel, der als Vater der modernen Genetik bezeichnet wird (Abb. 1), verfolgte ein ganz anderes Konzept. Dieses ist weitgehend übersehen worden, erweist sich aber gerade angesichts zahlreicher Befunde über sehr schnelle Artbildungsprozesse als ausgesprochen aktuell und fruchtbar. Es beruht auf präexistenter Vielfalt durch präexistente genetische Programme und ermöglicht Artbildung innerhalb weniger Generationen, da die Unterschiede zwischen den Arten nicht in langwierigen Mutations-Selektions-Zyklen aufgebaut werden müssen, sondern von Beginn an bereits angelegt sind. Sein Gesetz der Kombinationen von Merkmalsausprägungen erweist sich angesichts zahlreicher neuerer Befunde als ausgesprochen nützlich zum Verständnis der Veränderung von Arten innerhalb genetischer Familien*. Mendels Konzept – Mendel’sche Artbildung – soll in diesem Beitrag und in zwei Folgebeiträgen vorgestellt und Darwins Ansatz gegenübergestellt werden.
Kompakt
Die meisten Tier- und Pflanzenfamilien weisen eine enorme Vielfalt unterschiedlich angepasster Arten auf. Woher kommt diese Vielfalt? Gregor Mendels bahnbrechende wissenschaftliche Abhandlung aus dem Jahre 1866, Versuche über Pflanzenhybriden, bietet für diese Frage eine heute weitgehend übersehene Erklärung. In ihr beschreibt er Eigenschaften von Hybriden*, die sowohl dominante* als auch rezessive* Merkmalsausprägungen* haben, und legt dar, dass rezessive Merkmalsausprägungen in den Hybriden latent* vorhanden sind. Er beschreibt, wie diese latenten Merkmalsausprägungen zum Vorschein kommen und wie dadurch neue Sorten und Arten von Hülsenfrüchten erzeugt werden können. Mendels Entdeckungen lassen die genetische Strategie hinter der Entstehung der Arten (innerhalb von separaten Familien) erkennen. Sein Gesetz der Kombinationen von Merkmalsausprägungen resultiert aus Rekombination* der Merkmalsausprägungen bei der Meiose (Bildung der Geschlechtszellen).
Eigenartigerweise wird dieses Mendel‘sche Gesetz von Biologen meistens übersehen oder bewusst ignoriert. Dabei lässt sich mithilfe dieses Gesetzes die Entstehung einer großen Anzahl von phänotypischen* Variationen innerhalb genetischer Familien* zufriedenstellend erklären. Dies wiederum ist eine ausreichende Basis für die Entstehung von Arten innerhalb genetischer Familien (Grundtypen). Vielfalt innerhalb genetischer Familien und Prozesse der Artbildung (Aufspaltung von Arten) beruhen demnach auf bereits vorliegenden, präexistenten genetischen Anlagen. Wenn alternative Kombinationen dieser präexistenten Merkmalsausprägungen in den Nachkommen fixiert werden, kann daraus eine Fülle von neuen Arten und Gattungen entstehen. Die Summe aller dieser Kombinationen stellt die Grenzen dar, innerhalb derer eine Familie sich durch Ausprägung der angelegten Möglichkeiten entfalten kann.
Mendels Erkenntnisse können aus heutiger Sicht als Gegenentwurf zu der auf Charles Darwin zurückgehenden Theorie verstanden werden, wonach die Vielfalt der Arten (auch über genetische Familien hinaus) durch zufällige Mutationen* und deren langwierige Fixierung durch natürliche Auslese und andere Prozesse entstanden sind.
In diesem ersten Teil der auf drei Teile angelegten Artikelserie über Mendel’sche Artbildung werden Mendels Gesetz der Kombinationen von Merkmalsausprägungen und andere von ihm entdeckte Gesetzmäßigkeiten vorgestellt und erläutert, wie es mithilfe dieser Gesetze gelingt, die Verhältnisse beobachteter Merkmalsausprägungen bei Hybriden zu erklären. Dieses Gesetz ist Grundlage für das Verständnis der Entstehung einer großen Vielfalt von Merkmalsausprägungen und darauf aufbauender Artbildung.
Mendels Gesetze – verschiedene Formulierungen
Mendels Gesetze werden ihm zugeschrieben. Mendel hat sie jedoch nie formuliert, und ohne eindeutige Quelle wurde ihr Inhalt im Laufe der Zeit unterschiedlich wiedergegeben. Lehrbücher über Genetik auf Englisch haben die folgenden zwei (drei) Gesetze. Mendels erstes Gesetz „The Law of Segregation“ (Gesetz der Auftrennung) besagt, dass Merkmalszustände (Englisch: traits) (oder Allele) nach dem Zufallsprinzip an die nächste Generation bei der Meiose weitergegeben werden. Mendels zweites Gesetz „The Law of Independent Assortment“ (Gesetz der unabhängigen Auftrennung) besagt, dass Merkmale (Englisch: characters) (oder Gene) zufällig an die nächste Generation bei der Meiose weitergegeben (sortiert) werden (bei Kopplung von Genen am gleichen Chromosom kann diese Unabhängigkeit bzw. Zufälligkeit gestört sein). Mendels drittes Gesetz „The Law of Dominance“ (Gesetz der Dominanz) wird fast nie erwähnt und besagt, dass dominante Merkmalszustände (Allele) rezessive Merkmalszustände (Allele) maskieren (bzw. verbergen).
Lehrbücher über Genetik auf Deutsch nennen folgende drei Regeln (oder Gesetze). Regel 1, die „Uniformitätsregel“, besagt, dass Eltern, die unterschiedlich homozygot sind, einheitliche heterozygote (mischerbige) Nachkommen hervorbringen. Regel 2, die „Spaltungsregel“, besagt, dass Eltern, die in gleicher Weise heterozygot sind, sich beim Nachwuchs sowohl phänotypisch als auch genotypisch aufspalten. Regel 3, die „Unabhängigkeitsregel“, entspricht dem Gesetz der unabhängigen Auftrennung.
Woher stammt die Vielfalt in der Natur?
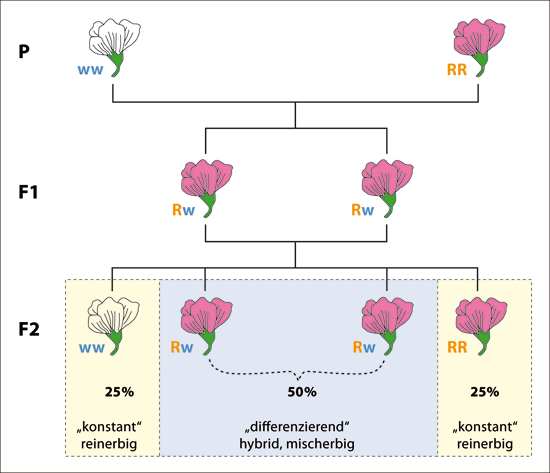
Abb. 2: Veranschaulichung der Uniformitätsregel (bei Generation F1) und der Spaltungsregel (bei Generation F2) bei einem monohybriden* dominant-rezessiven Erbgang.
Im unteren Teil der Abbildung wird gezeigt, was mit konstanten (reinerbigen, homozygoten) und differenzierenden (hybriden, mischerbigen, heterozygoten) Merkmalen gemeint ist.
Obwohl Mendel das Gesetz der Kombinationen von Merkmalsausprägungen entdeckt hat, das den Ursprung biologischer Vielfalt erklärt, und obwohl dieses den einzigen nachgewiesenen Mechanismus für den Ursprung eindeutiger Arten innerhalb eukaryotischer* Familien darstellt und obwohl Mendel das Rekombinationsgesetz mindestens dreimal in seinem berühmten Fachartikel über Pflanzenhybriden, der die Wissenschaftsdisziplin der Genetik ins Leben rief, nachgewiesen und formuliert hat, wird dieses sein Gesetz unbegreiflicherweise im Großen und Ganzen nach wie vor übersehen. Biologie- und Genetik-Lehrbücher schreiben Mendel zwei bzw. drei andere Gesetze zu: die sogenannte Uniformitätsregel, Spaltungsregel (Abb. 2) und Unabhängigkeitsregel. Mit unseren heutigen Erkenntnissen können wir diese Regeln als Gesetz über die Auftrennung der Allele* (Regel 1 und 2) und als Gesetz der unabhängigen Sortierung der Gene (Regel 3) bezeichnen (wie es in den meisten englischen Genetik-Lehrbücher steht; vgl. Kasten). Durch seine Forschung wurde die Gültigkeit dieser beiden Gesetze zweifellos nachgewiesen (abgesehen von der später entdeckten Möglichkeit der Kopplung von Genen), aber ausdrücklich formuliert hat er sie nie.
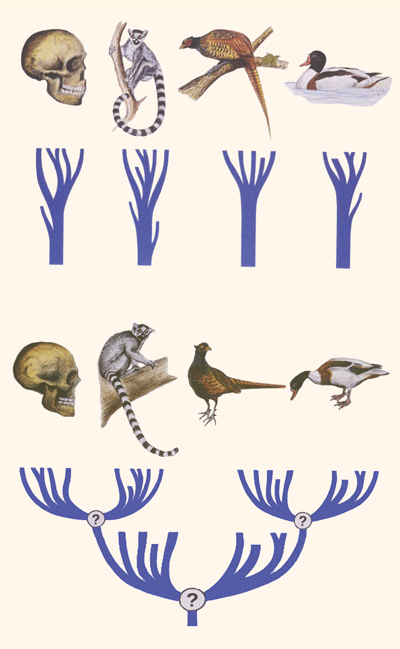
Abb. 3: Cis-Evolution versus trans-Evolution.
Bei der cis-Evolution (oben) sind mehrere separate Familien zu sehen. Jede Familie hat die Form eines Baumes.
Die Äste innerhalb eines Baumes werden durch Fortpflanzungsisolation auseinandergehalten, aber können durch sporadische Hybridisierung gelegentlich wiedervereint werden. Es gibt hier einen „Wald“ des Lebens. Dieser soll durch präexistente genetische Programme und Mendel’sche Artbildung zustande gekommen sein.
Bei der trans-Evolution (unten) sind dagegen alle Familien abstammungsmäßig miteinander verbunden. Es gibt hier nur einen einzigen großen Lebensbaum. Dieser soll durch Mutationen und Selektion zustande gekommen sein. (Nach Junker & Scherer 2013, verändert)
Wenn man sein Manuskript gründlich liest, stellt man fest, dass es zweifellos Mendels Ziel war, seine Leser auf das Gesetz der Kombinationen von Merkmalsausprägungen (d. h. die exponentielle Zunahme der Kombinierbarkeit der Merkmalausprägungen mit der einfachen Zunahme der Merkmale) aufmerksam zu machen. In einer ausgezeichneten neueren Besprechung des Mendel’schen Vermächtnisses (Ellis et al. 2011) wurde auf diese ungewöhnliche Situation hingewiesen. Darin wird die irreführende englische Übersetzung des Titels der Mendel’schen Abhandlung beklagt und bestätigt, dass Mendels Schwerpunkt auf der Speziation (Artbildung) lag, der Verwandlung von einer Art in eine andere. Sein Gesetz der Kombinationen von Merkmalsausprägungen erklärt, wie eine große Menge phänotypischer* Information auf elegante Weise in den Genomen* der Organismen codiert ist. Das ist nicht nur völlig ausreichend, um das breite Spektrum der beobachteten Vielfalt innerhalb eukaryotischer Familien zu erklären, sondern sein Vielfaltspotenzial geht weit darüber hinaus. Mendel gebührt volle Anerkennung für die Entdeckung des Gesetzes von der freien Kombinierbarkeit der Merkmale und für die Entdeckung des Mechanismus, der der Entstehung der Arten innerhalb genetischer Familien zugrunde liegt.1
Woher stammt die Vielfalt in der Natur? Die Entstehung der Arten ist ein faszinierendes Thema. Warum sind Leoparden und Jaguare einander so ähnlich und bleiben trotzdem immer verschieden? Warum sind Wölfe als eigene Art relativ homogen und doch stellen die Hunde, ihre künstlich gezüchteten Nachkommen, einen solchen Reichtum an unterschiedlichen Rassen dar? Warum sind einige Organismen in der Lage, eine große Vielfalt neuer Arten zu erzeugen, wenn sie Meeresinseln besiedeln, während andere das nicht können? Vor Darwin begnügte sich die Mehrheit in der Gesellschaft damit, biologische Vielfalt göttlichem Handeln zuzuschreiben, oder fand sich einfach damit ab, dass man sie eben nicht erklären kann. Aber nach der Veröffentlichung seines Buches suchte man nach einer Erklärung, die wissenschaftlichen Ansprüchen genügte.
Der Augustinermönch Gregor Mendel (Abb. 1) veröffentlichte die Lösung, doch obwohl sie klar formuliert war, wurden seine Entdeckungen von einer sich um sich selbst kreisenden wissenschaftlichen Gesellschaft missachtet und fielen in einen jahrzehntelangen Dornröschenschlaf. Dann aber wurde plötzlich sein Werk anerkannt, und zwar in drei voneinander unabhängigen Artikeln in den Ausgaben vom März, April und Juni 1900 des „Berichtes der Deutschen Botanischen Gesellschaft“, deren Autoren de Vries, Correns und von Tschermak waren. Correns machte deutlich: „Dass dies Verhalten von Bedeutung für die Frage ist, ob aus Bastarden (=Hybriden) Arten werden können, braucht wohl kaum bemerkt zu werden.“ Mendel hatte die Quelle für die Vielfalt in der Natur entdeckt. Später sollte man den zugrunde liegenden Vorgang als meiotische* Rekombination bezeichnen, und mit dieser Entdeckung hatte Mendel das Geheimnis der Entstehung der Arten gelüftet.
Die eukaryotische Welt ist äußerst vielfältig und umfasst Millionen von Organismenarten. Man könnte sie durch einen einzelnen Abstammungsbaum des Lebens darstellen (dann wäre sie monoarboreal) oder durch zahlreiche Abstammungsbäume, einen Abstammungswald des Lebens (dann wäre sie sylvan; vgl. Abb. 3). Beide Modelle gehen davon aus, dass ursprüngliche Arten zu zahlreichen von ihnen abstammenden Arten führen, aber sie haben ganz verschiedene Antworten auf das „Wie“ und das Ausmaß der Veränderungen. Mendels Abhandlung beschrieb den Mechanismus der Artbildung. Sie behandelte speziell die wohl wichtigste Frage in der Biologie: Sind Arten innerhalb bestimmter Grenzen fix, über die hinaus sie sich nicht ändern können (womit das Leben als Abstammungswald beschrieben würde), oder können Arten grenzenlos variieren (womit die Lebewesen in einem einzigen Abstammungsbaum dargestellt werden könnten)?
Natürlich benutzte Mendel Begriffe, mit denen wir heute meistens nicht so vertraut sind. Mit der Abfassung seiner Abhandlung erlebte die Genetik als Wissenschaftsdisziplin ihre Geburtsstunde. Während Mendel seine Experimente in den Jahren zwischen 1854 und 1865 durchführte, erschien schließlich Darwins Über die Entstehung der Arten durch natürliche Zuchtwahl (1859). Mendel besaß sein eigenes Exemplar der deutschen Übersetzung dieses Buches aus dem Jahre 1863 (Mawer 2006). Darwin verstand das Leben als einen einzigen Abstammungsbaum. Da Mendel begriff, was die Ergebnisse seiner Experimente bedeuteten, neigte er eher dazu, die Natur als einen Abstammungswald aufzufassen. Ihn bedrückte jedoch die Tatsache, dass er seine Forschungen nur bei einigen Hülsenfrüchten durchgeführt hat. Mehrere Male empfahl er, dass seine Untersuchungen an anderen Organismen durchgeführt werden sollten, und er war deshalb eher vorsichtig mit Verallgemeinerungen.
Hinweise: Eine ausführlichere Darstellung der Mendel‘schen Artbildung ist veröffentlicht unter https://www.wort-und-wissen.de/artikel/sp/b-19-3_mendel.pdf
Mendels Originalarbeit ist unter mendel.pdf abrufbar.
Glossar
Allel: Zustandsform bzw. Variante ein und desselben Gens. Artbildung: Artaufspaltung; Prozess, durch den im Laufe von Generationen sich eine Art (Spezies) in zwei Tochterarten aufspaltet. dihybrid: in zwei Merkmalen mischerbig. dominant: Ein → Allel ist dominant, wenn es die → Merkmalsausprägung eines anderen (→ rezessiven) Allels unterdrückt. Epistasis: Gen-Wechselwirkung. Die Wirkung des einen (epistatischen) Gens verdeckt eines der Merkmale des anderen (hypostatischen) Gens. Statt erwarteter zweier Merkmale wird nur eines ausgebildet. Dominante Epistasis liegt vor, wenn die Anwesenheit eines einzelnen dominanten → Allels des epistatischen Gens eines der Merkmale des hypostatischen Gens verdeckt. Rezessive Epistasis liegt vor, wenn die Anwesenheit beider rezessiver Allele des epistatischen Gens die Ausprägung eines oder mehrerer Merkmale des hypostatischen Gens unterdrückt. eukaryotische Organismen: Lebewesen mit echten Zellkernen. Sie sind normalerweise zur Meiose fähig. Expression (Genexpression): Das Ablesen und somit Nutzbarmachen von Genen bzw. der durch sie codierten Information im Zellbetrieb. exprimieren: → Expression. genetische Familie: Familie, deren zugehörende Arten durch verschiedene Merkmalskombinationen einer präexistenten Vielfalt charakterisierbar sind. Genom: Das gesamte Erbgut eines Individuums Genotyp: Gesamtheit der Erbfaktoren. heterozygot: mischerbig (das Gen liegt in zwei verschiedenen Allelen vor). homozygot: gleicherbig (das Gen liegt in zwei gleichen Allelen vor). Hybrid: Mischling. Isolation: Trennung von Populationen und Unterdrückung eines Genaustauschs. latent: versteckt, verborgen (hier: im Erbgut). Ein → Merkmal oder → Merkmalszustand ist latent, wenn es / er zwar genetisch angelegt ist, aber nicht ausgeprägt (→ exprimiert) wird. Meiose: Reifeteilung, Bildung der Geschlechtszellen. Merkmal: Bestimmtes Kennzeichen des Phänotyps, entspricht einem Gen. Beispiel: Das Merkmal „Blütenfarbe“ (nicht: die tatsächlich ausgeprägte Farbe selbst). Merkmalsausprägung: spezifische phänotypische Ausprägung eines → Merkmals, entspricht einem bestimmten → Allel eines Gens. Beispiel: Verschiedene Farben einer Blüte. monohybrid: in einem Merkmal mischerbig. Mutation: Spontane oder künstlich ausgelöste Änderung des Erbguts. Phänotyp, phänotypisch: äußere Erscheinungsform, die Morphologie, Anatomie oder Physiologie betreffend. Rekombination: Durchmischung des Erbgutes bei der sexuellen Fortpflanzung. rezessiv: Ein → Allel ist rezessiv, wenn es durch die → Merkmalsausprägung eines anderen (→ dominanten) Allels unterdrückt wird. trihybrid: in drei Merkmalen mischerbig. Zygote: Zelle von → Eukaryoten, die bei der geschlechtlichen Fortpflanzung durch Verschmelzung von einer Eizelle und einem Spermium entsteht.
Mendels Gesetz der Kombinationen von Merkmalsausprägungen (meiotische Rekombination)
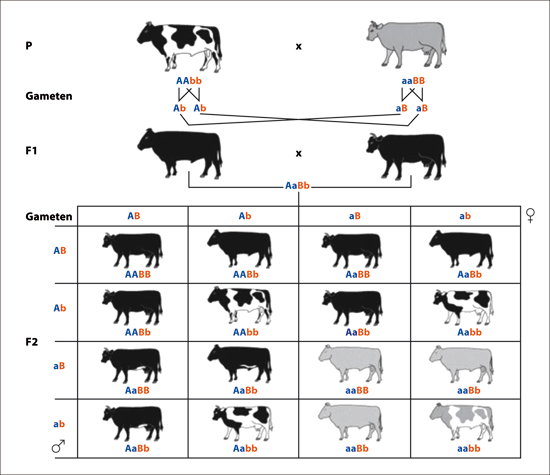
Abb. 4: Veranschaulichung der Uniformitätsregel (bei Generation F1) und der Spaltungsregel (bei Generation F2) bei einem dihybriden dominant-rezessiven Erbgang. Einfarbig ist hier dominant über gescheckt; schwarz ist dominant über grau. In der F1-Generation sind die rezessiven Allele (für grau und gescheckt) verdeckt (latent).
Darwin war ein Zeitgenosse Mendels und ein leidenschaftlicher Biologe, der viele weitreichende Entdeckungen machte. Schon aus dem Titel seines berühmten Buches geht eindeutig der Mechanismus hervor, den er für den Ursprung der Arten verantwortlich machte. Der volle Titel lautete nämlich: „Über die Entstehung der Arten durch natürliche Zuchtwahl oder die Erhaltung der begünstigten Rassen im Kampfe ums Dasein“. Darwin glaubte, dass die natürliche Zuchtwahl das Mittel sei, durch das neue Arten ins Dasein gelangen. Heute wird ihm zugeschrieben, er habe das Geheimnis der Artbildung gelüftet. Jedenfalls wurde durch seinen Vorschlag der Mechanismus von der göttlichen Ebene auf die irdische Ebene verschoben. Eigentlich ist seine Prämisse jedoch nicht richtig. Selektion (=Zuchtwahl) ist kein Mechanismus, der die Entstehung neuer Merkmalsausprägungen oder neuer Arten verursacht. Sie begünstigt jedoch deren Überleben. Im Gegensatz dazu wies Mendel anhand seiner Untersuchungen von Pflanzenhybriden nach, wie neuartige Merkmalsausprägungen wirklich entstehen. Die meiotische Rekombination ist die notwendige und auch ausreichende Quelle von neuartigen Merkmalsausprägungen.2 Darüber hinaus bietet dieser Mechanismus eine einfache Erklärung, wie neue Arten entstehen. Man hat noch keinen anderen Mechanismus nachgewiesen, der entweder notwendig oder ausreichend wäre, um die Entstehung neuer (nicht-trivialer) Arten bei Eukaryoten zu erklären. (Wie dieser Mechanismus funktioniert, ist Gegenstand der beiden Folgeartikel.) Nicht-meiotische Organismen wie Prokaryoten verwenden andere Mechanismen, um genetische Information auszutauschen.
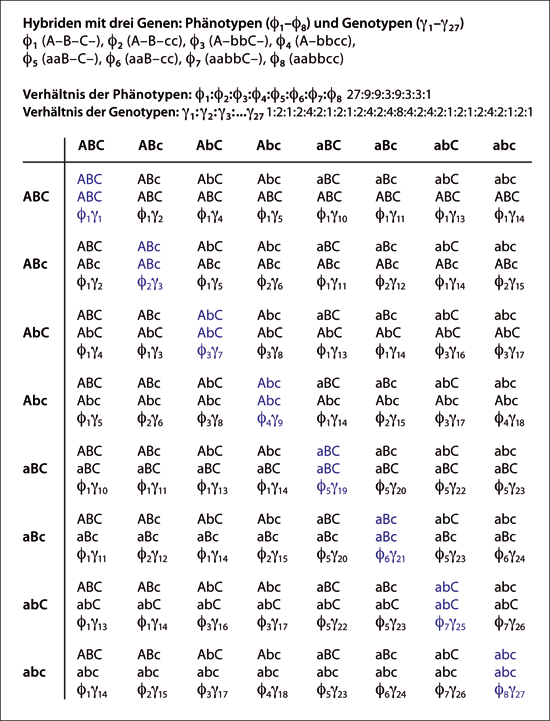
Abb. 5: Mendel‘sche Erzeugung von Vielfalt. Punnett-Quadrat mit drei Genen, das die möglichen Ergebnisse einer trihybriden Selbstkreuzung zeigt. Keines der Gene ist mit einem anderen verknüpft. Aus den drei heterozygoten Genen entstehen 8 (23) einzigartige haploide Gameten, die am oberen und linken Rand aufgelistet sind. Wenn sich diese bei der Befruchtung kombinieren, entstehen 8 (23) einzigartige Phänotypen, 27 (33) einzigartige Genotypen und 64 (43) einzigartige diploide Kombinationen von Allelen. Es gibt so viele einzigartige Gameten, wie es einzigartige Phänotypen gibt. Die 8 Phänotypen (f1-f8), ihre redundanten Genotyp-Bezeichnungen (ein Bindestrich zeigt an, dass jedes der beiden Allele vorhanden sein kann) und ihr erwartetes Verhältnis sind oberhalb der Matrix aufgeführt. Die Leitdiagonale (von oben links nach unten rechts, kursiv blau) zeigt alle 8 (23) konstanten, reinerbigen, panhomozygoten Genotypen. Wenn sie unter reproduktiver Isolation gehalten werden, handelt es sich um neuartige Sorten bzw. Spezies. Die Rückendiagonale (von oben rechts nach unten links) zeigt den pan-heterozygoten, panhybriden Genotyp. Dieser besitzt die maximale Merkmalsinformation. Bei zwei (oder mehr) Stamm-Individuen können jedoch doppelte (oder mehr) Merkmalsinformationen übertragen werden. Das erwartete Verhältnis der 27 Genotypen (g1-g27) ist ebenfalls angegeben. Die 8 Phänotypen (f1-f8) und 27 Genotypen (g1-g27) sind in der Matrix angegeben.
Das mögliche Ausmaß der Variation
Wie viel Variation kann man vernünftigerweise auf der Grundlage der Mendel‘schen Experimente mit sieben Merkmalen bei Erbsen erklären, mit denen Mendel experimentiert hat? Es hat sich gezeigt, dass drei Prinzipien bei der Mendel’schen Artbildung wirksam sind: latent* vorhandene Information, das Gesetz der Kombinationen von Merkmalsausprägungen* und der Verlust der Mischerbigkeit. Das erste Prinzip beobachtete Mendel bei seinen frühesten Experimenten mit Hybriden. Ihre Merkmale wiesen dominante und rezessive Ausprägungen auf. Mendel bezeichnete Merkmalsausprägungen, die latent werden, als rezessiv, „weil die damit benannten Merkmale an den Hybriden zurücktreten oder ganz verschwinden.“ Solche latente genetische Information ist für die Artbildung entscheidend. Diese Information ist von Anfang an vorhanden. Sie muss nie erzeugt werden und ist immer potenziell verfügbar. Genetische Information wird durch mindestens drei Mechanismen in latentem Zustand gehalten: Dominanz, Epistasis* und Transposition (Näheres dazu im Teil 2 der Artikelserie).
Das zweite Prinzip, das Mendel bei seinen Experimenten mit Hybriden beobachtete, war das Gesetz der Kombinationen von Merkmalsausprägungen. Zwar hat Mendel das Gesetz ausdrücklich formuliert und in seiner Abhandlung dreimal angeführt, doch hat er nie eine Anerkennung dafür erhalten. Stattdessen werden ihm zwei (oder drei) andere Gesetze zugeschrieben, die er nicht speziell formuliert hat. Wenn man den gesamten Sachverhalt berücksichtigt, sollte das Gesetz der Kombinationen von Merkmalsausprägungen eigentlich als sein erstes Gesetz gelten und die anderen könnten sein zweites und drittes Gesetz darstellen. Mit Hilfe des Gesetzes der Kombinationen von Merkmalsausprägungen gelang es Mendel, das zu erwartende Verhältnis von Genotypen* bei monohybriden*, dihybriden* und trihybriden* Kreuzungen vorherzusagen. Er entdeckte das Phänotypenverhältnis von 3:1 bei dominanten zu rezessiven Merkmalsausprägungen (Abb. 2). Doch Mendels Forschung ging noch weiter in die Tiefe. Er wusste, dass der dominante Phänotyp sowohl konstante (homozygote) als auch differenzierende (heterozygote) Merkmale enthält (zu diesen Begriffen siehe Abb. 2). Er zeigte auf, dass bei Letzteren das monohybride (1 Gen) Genotypenverhältnis tatsächlich 2:1:1 beträgt. Dann wies er nach, dass das dihybride (2 Gene) Genotypenverhältnis bei 9 einzigartigen Genotypen 1:1:1:1:2:2:2:2:4 lautet (Abb. 4). In Genetik-Lehrbüchern wird das Verhältnis mit 9:3:3:1 angegeben, weil die gleich gestalteten Phänotypen normalerweise zusammengefasst werden, auch wenn sie unterschiedliche Genotypen besitzen. Mendel wies sogar nach, dass das trihybride (3 Gene) Genotypenverhältnis von 27 einzelnen Genotypen 1:1:1:1:1:1:1:1:2:2:2:2:2:2:2:2:2:2:2:2:4:4:4:4:4:4:8 beträgt (siehe Abb. 5).
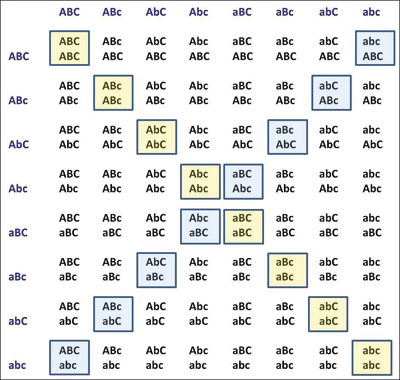
Abb. 6: Punnett-Quadrat mit 3 Merkmalen in vereinfachter Darstellung; es sind nur Genotypen angezeigt. Von den 64 Kombinationsmöglichkeiten sind acht (Leitdiagonale) für alle Merkmale reinerbig (konstant); vgl. Abb. 5. Zur Erläuterung der unterlegten Diagonalen siehe Text zu Abb. 5.
Der britische Biologe Reginald Punnett (1911) veröffentlichte eine effektive Methode, mit der er die Vielfalt und Häufigkeit der Nachkommen bei solchen Experimenten darstellen konnte. Dabei benutzte er ein „Schachbrett“, das heutzutage nach ihm benannte Punnett-Quadrat. Abb. 5 und 6 zeigen Punnett-Quadrate für drei Merkmale (bzw. Gene), welche trihybride (3 Gene) Eigenkreuzungen darstellen. Das Punnett-Quadrat mit drei Merkmale (Genen) zeigt die 27 verschiedenen Genotypen, von denen Mendel berichtet hat. Es zeigt auch, dass diese 27 Genotypen in acht verschiedene Kategorien von Phänotypen eingeordnet werden können. Mendels „konstante“ und „differenzierende“ Pflanzen werden heutzutage als „reinerbige“ und „hybride“ Pflanzen bezeichnet. Die Hybride mit drei Merkmalen bringt acht einzigartige „reinerbige“ Nachkommen (acht konstante oder unveränderliche Arten) hervor, wenn alle drei Gene homozygot werden. Diese bilden die Hauptdiagonale des Punnett-Quadrats.
Die einzelnen Elemente der Trihybride (mit den acht unterschiedlichen Phänotypen) sehen zwar alle gleich aus, enthalten aber doch eine unterschiedliche Anzahl von heterozygoten Genen und wären von Mendel zumeist als „differenzierend“ eingeteilt worden. Dennoch kommt jeder Phänotyp immer einmal in einer konstanten (reinerbigen) Form vor. Mendel wies nach, dass hybride Organismen dem Gesetz der Kombinationen von Merkmalsausprägungen folgen. Deshalb steigt die Vielfalt exponentiell mit der Zahl der Merkmale (Gene). Eine Hybride mit „n“ Merkmalen (oder Genen) kann 2n verschiedene konstante Nachkommen und 3n verschiedene Genotypen erzeugen. Sie kann auch 2n verschiedene Phänotypen erzeugen. Des Weiteren untersuchte Mendel bei Erbsen insgesamt sieben differenzierende Merkmale und er beschrieb, dass er alle 27 (128) einzigartigen Phänotypen beobachtet hat. Da sein Gesetz von Natur aus exponentiell ist, erklärt es leicht die Entstehung einer Vielzahl von Phänotypen: 10 differenzierende Merkmale (heterozygote Gene) erzeugen bereits etwa tausend verschiedene reinerbige Phänotypen, 20 differenzierende Merkmale (heterozygote Gene) ca. eine Million verschiedene reinerbige Phänotypen. Hier haben die Variation der Arten und die Vielfalt in der Natur letztendlich ihren Ursprung.
Ausblick: Evolution eines Grundtyps mit neun Merkmalen
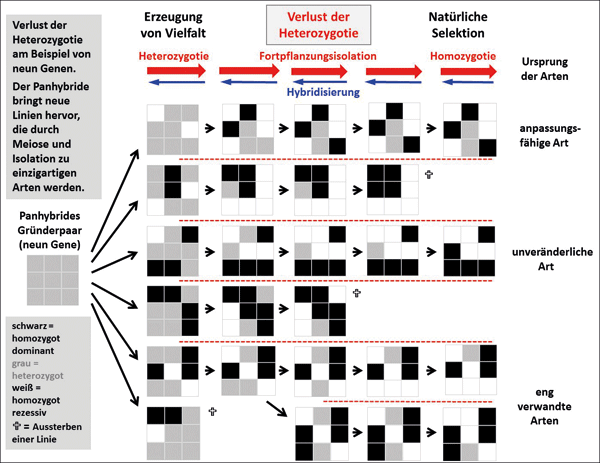
Abb. 7: Evolution eines Grundtyps mit neun Merkmalen. Erläuterungen im Text.
Nebenstehende Abbildung verdeutlicht, wie aus einem panheterozygoten Gründerpaar, bei dem alle Merkmale mischerbig (heterozygot) sind, Vielfalt ausgeprägt werden kann und verschiedene Arten entstehen können.
Genotypen* der Population (bzw. Art) werden als 3x3-Matrizen dargestellt. Jede Spalte mit 3x3-Matrizen repräsentiert eine aufeinanderfolgende Generation und jede Zeile die evolutionäre Linie einer Population (bzw. Art). Jede Zelle in einer Matrix repräsentiert ein Merkmal: schwarz, homozygot dominant; grau, heterozygot; weiß, homozygot rezessiv. Jede graue Zelle zeigt das dominante (schwarze) Merkmal im Phänotyp, kann aber ein rezessives (weißes) Allel an die nächste Generation weitergeben. Die Entwicklung von Merkmalen ist ein stochastischer Prozess. Nachdem die Populationen isoliert wurden (gestrichelte Linien), variieren nur heterozygote Merkmale (grau). Die Erzeugung von Vielfalt, der Verlust von Heterozygotie und die natürliche Selektion sind laufende Prozesse. Der Verlust der Heterozygotie führt zu einzigartigen Konstellationen homozygoter dominanter und rezessiver Merkmale, d.h. zum Ursprung der Arten. Populationen (Arten) sterben aus verschiedenen Gründen aus (?). Die letzte Spalte stellt die heutigen Populationen (Arten) dar. Die meisten haben eine gewisse Heterozygotie und das Potenzial zur Diversifizierung bewahrt. Eine Art (3. Reihe) ist völlig homozygot und „unveränderlich“. Die Hybridisierung kann den Verlust der Heterozygotie (Pfeilkette) teilweise umkehren. Die Hybridisierung der oberen überlebenden Population (Art) mit der „unveränderlichen“ Population (Art) stellt den ursprünglichen Phänotyp der oberen Linie wieder her, aber Heterozygotie ist nur bei den Merkmalen 2, 3, 5, 7 und 8 gegeben. In den beiden untersten Linien ist ein Verzweigungsereignis aufgetreten und die vorhandenen Nachkommen sind eng miteinander verwandt. Die Hybridisierung zwischen den beiden stellt den Phänotyp ihres jüngsten gemeinsamen Vorfahren wieder her, aber Heterozygotie ist nur bei den Merkmalen 2, 7 und 8 gegeben. Kein Hybridisierungsereignis zwischen bestehenden Populationen (Arten) kann das dominante Merkmal von Merkmal 1 oder die rezessive Ausprägung von Merkmal 4 wiederherstellen. Diese Information ist aus der Population verloren gegangen.
In den Teilen 2 und 3 dieser Artikelserie werden diese Vorgänge genauer beschrieben und anhand von Beispielen erläutert.
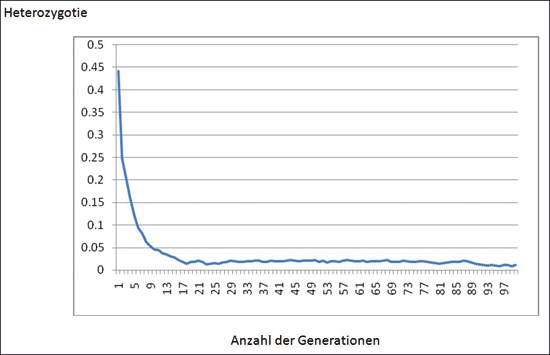
Abb. 8: Anteil heterozygoter Gene, aufgetragen gegen die Anzahl der Generationen in einer typischen Population (0,5 = 50 %). Verlust der Heterozygotie, was einer kompletten adaptiven Radiation einer genetischen Familie mit vielen neuen Arten und Gattungen entspricht, braucht weniger als zwanzig Generationen.
Das dritte von Mendel beschriebene Prinzip ist der Trend zur Konstanz (= Verlust der Heterozygotie). Das ist für die Artbildung entscheidend, denn nach diesem Prinzip werden heterozygote Gene in „fixierte“ homozygote Gene umgewandelt. Mendel war mit den Arbeiten einiger anderer Pflanzenzüchter wie Gärtner und Kölreuter vertraut. Sie hatten beobachtet, dass Hybriden die Tendenz haben, zu den Elternformen (reinerbigen Formen) zurückzukehren. Anhand eines Beispiels mit einem Merkmal (Gen) erläuterte Mendel, wie entsprechend seinem Gesetz eine Hybride (Heterozygote) innerhalb von zehn Generationen zur Entstehung von gleichen Anzahlen beider konstanten (homozygoten) Elternformen führt und fast keine Hybriden mehr übrig bleiben (weniger als eine von tausend: Verlust der Heterozygotie). Mendel beschreibt klar das Prinzip des Verlustes der Heterozygotie.
Wie schnell tritt der Verlust von Heterozygotie in der Population auf? Dauert es Hunderte von Generationen? Mendel beobachtete, dass eine heterozygote Population innerhalb von 10 Generationen nahezu 50 % homozygot dominant und 50 % homozygot rezessiv geworden war. Buri (1956) untersuchte 107 Populationen von heterozygoten Fruchtfliegen. Nach der Generation 6 erschienen bereits Populationen, die zu 100 % homozygot waren. Mit der Generation 19 waren fast alle Populationen entweder 100 % homozygot dominant oder 100 % homozygot rezessiv. Populationen mit vielen heterozygoten Genen zeigen einen ebenso schnellen Verlust der Heterozygotie. Mathematische Modelle von Populationen mit 25 heterozygoten Genen ergaben in ähnlicher Weise, dass nur 19 Generationen benötigt wurden, damit fast alle Gene entweder homozygot dominant oder rezessiv werden (vgl. Abb. 8).
Ausblick
In den beiden folgenden Teilen dieser Serie über Mendel‘sche Artbildung wird es um die Frage gehen, durch welche Mechanismen ein enormes Ausmaß an Variation schon vorhanden (präexistent) sein kann, aber zunächst – im heterozygoten Zustand – in einem latenten (verborgenen) Zustand verbleibt. Anschließend wird erklärt, wie diese latente Information ausgeprägt wird und wie im Laufe weniger Generationen eine Aufspaltung in zahlreiche unterschiedliche Arten erfolgen kann. Es zeigt sich: Es gibt einen nachvollziehbaren Mechanismus rascher Artbildung auf der Basis präexistenter Information und dieser Ansatz erklärt die heute vorliegenden Befunde über schnelle Artbildung besser als das Darwin‘sche Konzept.
1 Der hier verwendete Begriff „Entstehung der Arten“ ist weder mit dem allgemeinen Ursprung des Lebens noch mit der Entwicklung eines einzigen Lebensbaums vom bakterienähnlichen Organismus zum Menschen gleichzusetzen (wie weiter unten erläutert). Die Entstehung der Arten bezieht sich hier auf die Bildung von Arten, wie sie nach wissenschaftlich anerkannten Regeln beschrieben werden, ausgehend von einem eng verwandten gemeinsamen Vorfahren, meist innerhalb einer eukaryotischen (meiotischen) Familie. Familien sind zum Beispiel die Equidae (Pferdefamilie) oder die Felidae (Katzenfamilie).
2 Zumindest für Eukaryoten bzw. meiotische Organismen (oft mit sexueller Fortpflanzung assoziiert).
Gene frequency drift in small populations of mutant Drosophila. Evolution 10, 367–402.
On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life. John Murray, London.
Mendel, 150 years on. Trends Plant Sci. 16, 590-596.
Evolution – ein kritisches Lehrbuch. Gießen, 7. Aufl.
Gregor Mendel. Harry N. Abrams, inc. New York, N.Y.
Versuche über Pflanzenhybriden. Verh. Naturforsch. Vereins Brünn 4, 3-47.
Mendelism, 3rd ed. MacMillan Co. New York, NY.
Themen | Kurzbeiträge | Streiflichter
Studiengemeinschaft WORT und WISSEN e.V.
Letzte Änderung: 6/21/21
Webmaster