Themen • Kurzbeiträge • Streiflichter
„Schildkröten-Nashorn“: Riesiger Konkurrent der Dinosaurier
von Henrik Ullrich
Studium Integrale Journal
26. Jahrgang / Heft 2 - Oktober 2019
Seite 109 - 112
Zusammenfassung: Der Fund eines elefantengroßen Tieres mit Namen Lisowicia bojani – ein naher Verwandter der Säugetiere – in Fundschichten der späten Trias in Polen sorgt für mehrere Überraschungen. Wieder einmal müssen bisherige Vorstellungen zum Lebensumfeld der Dinosaurier sowie zur Evolution der Säugetiere hinterfragt oder sogar aufgegeben werden.
• • • • • •
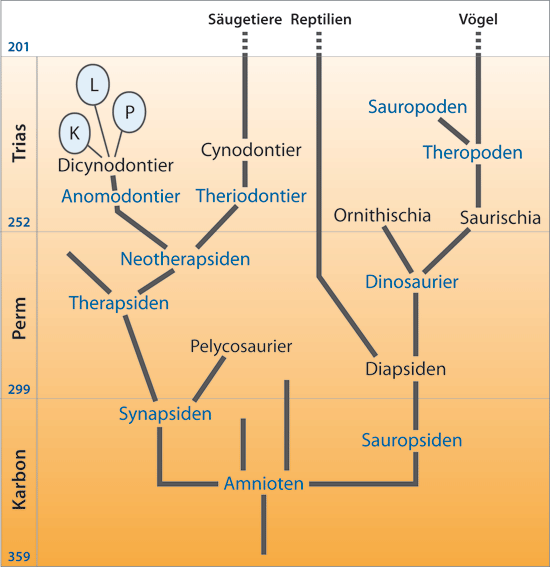
Abb. 1: Stark vereinfachchte Darstellung der Abstammungsverhältnisse der Amniota nach evolutionstheoretischen Deutungen, mit ungefährer Einordnung in die geologischen Formationen (links; MrJ = Millionen radiometrische Jahre).
Demnach kennzeichnen zwei große Entwicklungslinien die Amniota. Ein Zweig beinhaltet die Reptilien/Sauropsiden, zu denen der Großteil der fossilen und alle heutigen Echsen zählt. Der andere Zweig umfasst die Synapsiden, aus denen die Säugetiere hervorgegangen sein sollen. Es sind nicht alle Gruppen genannt, sondern die, welche für die Abstammungsverhältnisse besonders wichtig sind. Die im Text erwähnten Gruppen sind farblich hervorgehoben. K Kannemeyeria, L Lisowicia bojani, P Placerias. Dinosaurier und Säugetiere – ein systematischer Überblick
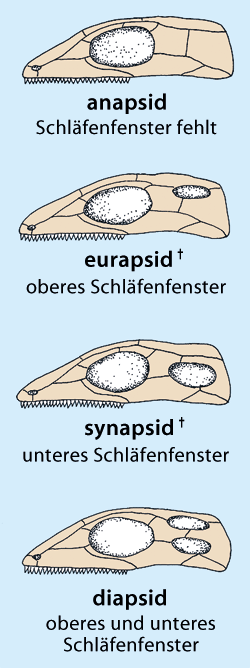
Abb. 2: Verschiedene Typen des Schläfenfensters bei Reptilien nach Romer & Parsons (1991). Der Typ des Schläfenfensters dient zur Klassifikation der Reptilien. Die säugerähnlichen Reptilien sind Synapsiden. (Aus Junker & Scherer 2013)
Unter den Evolutionsbiologen geht man davon aus, dass am Ende des Karbon vor ca. 300 MrJ (Millionen radiometrische Jahre) eine evolutionäre Aufspaltung der Amnioten1 in den Zweig der Sauropsiden2 (hypothetische Vorfahren u. a. der Dinosaurier, heutigen Reptilien und Vögel) und den der Synapsiden3 (hypothetische Vorfahren u. a. der Säugetiere) erfolgte (vgl. Abb. 1). Im Verlauf des Perm (ca. 299–252 MrJ) und der frühen Trias (ab ca. 252 MrJ) stellten dann, entsprechend der evolutionstheoretischen Interpretation, die Abkömmlinge der Synapsiden, die Therapsiden, die ökologisch dominierende Gruppe dar. Sie zeigten in diesem Überlieferungshorizont zunächst eine große Formenvielfalt. Das Aufkommen der Dinosaurier jedoch führte nach herrschender Ansicht zu einer Zurückdrängung der Therapsiden. Im Schatten der Riesenechsen sei es nur wenigen und kleinen Vertretern der Therapsiden gelungen, sich bis zum Ende der Trias (vor 201 MrJ) in einzelnen ökologischen Nischen zu behaupten. Darunter befanden sich unscheinbare, säugetierähnliche Formen, Eutherapsiden mit zwei Untergruppen, den Theriodontiern und Anomodontiern (vgl. Abb. 1 und Abb. 2). Zu den Theriodontiern gehörten u. a. die Cynodontier (Hundezähner), die als die unmittelbaren Vorfahren der Säugetiere angesehen werden.
Der hier vorgestellte Überraschungsfund, Lisowicia bojani, wird systematisch zu den Anomodontiern, speziell der Untergruppe der Dicynodontier (Zwei-Hundezähner), gestellt. Diese Seitengruppe ist erstmals in Sedimenten fossil überliefert, denen ein Alter von ca. 270 MrJ zugewiesen wurde. Die bisher letzten bekannten Vertreter dieser Gruppe sind neben den Lisowicia-Giganten, die in den jüngsten Schichten der Trias (208 MrJ) in Polen entdeckt wurden, Formen, welche aus der Unterkreide (ca. 110 MrJ) Australiens stammen (Thulborn et al. 2003).
Die Dicynodontier repräsentierten die artenreichste Gruppe innerhalb der Therapsiden. Fossilien der Dicynodontier fand man bislang in Afrika, Amerika, Australien und Asien. Frühe Dicynodontier waren nicht größer als 30 cm. Nur wenige Gattungen, so die Theorie, überlebten das vermutete große Aussterbeereignis am Ende des Perm (vor ca. 251 MrJ).
Aktuelle fossile Befunde fordern eine Neubewertung der o.g. Vorstellungen. Einige Vertreter der Dicynodontier durchliefen demnach im Verlauf der Trias, unbeeindruckt von oder gerade wegen der Existenz der Dinosaurier, eine überraschend schnelle Radiation mit dem Ergebnis, dass am Ende der Trias unerwartet große und vielgestaltige Formen wie Kannemeyeria mit einer Länge von etwa 3 Metern und Placerias, ein nilpferdähnlicher Dicynodontier mit ca. 3,5 Metern Länge und einem Gewicht von ungefähr einer Tonne, oder Lisowicia bojani auftraten. Letzterer erreichte nicht nur die Größe heutiger Elefanten, sondern auch deren Gewicht (Abb. 3 und 4). Zeitgleich mit den Dinosauriern – also in den entsprechenden Fundhorizonten der Trias – traten also diese Formen als deren ernstzunehmende Konkurrenten in einem gemeinsamen Lebensraum in Erscheinung.

Abb. 3: Der Dicynodontier Placerias mit rechtwinklig zum Körper angeordneten Oberarmknochen und senkrecht unter dem Körper stehenden Hinterextremitäten. (Rainbow Forest Museum, gemeinfrei) Ein überraschender Fund
Lisowicia war das bis dahin größte Landwirbeltier, welches nicht zu den Dino-sauriern gehörte und zeitgleich mit ihnen lebte.
Fossile Überreste von zwei Vertretern der Art Lisowicia bojani entdeckten Forscher im Süden Polens nahe der Ortschaft Lisowice. Die Fundschicht wird der Obertrias zugezählt und auf 208 MrJ datiert. Sulej & Niedzwiedzki (2018) vermuten, dass der 4,50 Meter lange, 2,60 Meter hohe und neun Tonnen schwere Gigant das bis dahin größte Landwirbeltier war, welches nicht zur Formengruppe der Dinosaurier gehörte und gleichzeitig mit diesen lebte.
Als typisches Kennzeichen der Dicynodontier (Zwei-Hundezähner) besaß Lisowicia bojani anstelle eines Gebisses einen Schnabel aus Horn (ähnlich dem heutiger Schildkröten), seitlich davon ragten zwei massive Zähne hervor, die Stoßzähnen glichen (Abb. 4).
Das Bild kann online nicht zur Verfügung gestellt werden.
Abb. 4: Größenvergleich von Lisowicia mit einem heutigen Elefanten. (Bild: Science/Tomasz Sulej und Grzegorz Niedzwiedzki)
Typisch für die übrigen Dicynodontier waren ihre rechtwinklig bzw. horizontal zum Körper angeordneten Oberarmknochen, während die hinteren Extremitäten senkrecht unter dem Körper standen (vgl. Abb. 3). Demgegenüber zeigt Lisowicia bojani auch senkrecht unter dem Körper angeordnete Oberarm- und Unterarmknochen. Dies ist eine anatomische Besonderheit, wie sie auch bei Säugetieren (z. B. bei Nilpferd, Rhinozeros) und Dinosauriern zu finden ist. Mit dieser Anordnung verband sich eine völlig andere Konfiguration und Physiologie des Muskelapparates als bei den übrigen Vertretern der Dicynodontier (z. B. Ursprungs- und Ansatzstellen der Muskulatur an den Knochen, was Bewegungsabläufe und Koordination betrifft). Ein weiteres physiologisches Merkmal, das Lisowicia bojani mit den Dinosauriern, jedoch nicht mit seinen nächsten Verwandten teilte, lag in dessen Fähigkeit, rasch und dauerhaft zu wachsen. Das konnten die Autoren aus der Struktur und dem Aufbau der Knochen ableiten.
In der fossilen Überlieferung tritt Lisowicia bojani wie die anderen erwähnten großen Formen Kannemeyeria und Placerias unvermittelt, d. h. ohne Zwischenformen, auf; mögliche Vorfahren werden in der Gruppe der Dicynodontier aus der frühen Trias vermutet (Sulej & Niedzwiedzki 2018). Somit eröffnet sich eine fossile Fundlücke von ca. 45 MrJ.
Neue Bewertung der Lebenswelten in der Trias und alte Fragen an die Modelle der Evolution
Die beeindruckenden Funde erfordern, so Sulej & Niedzwiedzki (2018), in verschiedener Hinsicht eine neue Bewertung der Lebenswelten in der Trias insbesondere bezüglich der morphologischen Vielfalt und Verbreitung der Dicynodontier und der bislang favorisierten evolutionstheoretischen Modelle über ihre Entstehung.
Vielfalt und Größe trotz der Dominanz der Dinosaurier
Die Entwicklung von Riesenwuchs bei Pflanzenfressern (Gigantismus) während der späten Trias (vgl. Abb. 1) war kein Alleinstellungsmerkmal der Dinosaurier, sondern ist auch bei anderen Tiergruppen in der späten Trias nachweisbar. Auch die Dicynodontier, insbesondere Lisowicia bojani, konnten riesige Größen erreichen. Die Theorie, dass säugetierähnliche Formen, die als Ur-Säugetiere bezeichnet werden, erst nach dem Verschwinden der Dinosaurier große bzw. gigantische Formen entwickeln konnten, ist damit nicht mehr haltbar. Sulej & Niedzwiedzki (2018) schließen, dass die sogenannten Ur-Säugetiere und andere Formen von Synapsiden gemeinsam mit Dinosauriern lebten, sie wurden nicht, wie bisher vermutet, von den Dinosauriern im Lauf der Trias verdrängt und zur Bedeutungslosigkeit verurteilt.
Fundort Europa – nicht-überlieferte Ökosysteme, nicht-überlieferte Fossilien
Auch der Fundort dieser riesigen Dicynodontier in Polen war eine Überraschung. Damit wurde nicht nur die erstaunliche morphologische Vielfalt am Ende der Trias innerhalb der Dicynodontier bestätigt, sondern auch eine unerwartet weite Verbreitung dieser Tiergruppe. Bislang vermutete man nur ein auf einzelne Regionen begrenztes Auftreten der Dicynodontier. Die älteren Fossilien von Dicynodontiern stammten vor allem aus Afrika, Asien und Amerika. Lediglich ein Unterkiefer und ein paar Knochen waren in Europa gefunden und den Dicynodontiern zugeordnet worden. Die Autoren vermuten als Erklärung für den nun bis Europa reichenden Lebensraum der Dicynodontier die Existenz des Superkontinents Pangaea am Ende der Trias, wo Erdteile noch miteinander über Landbrücken verbunden waren und den Dicynodontiern eine großflächige Ausbreitung ermöglichten.
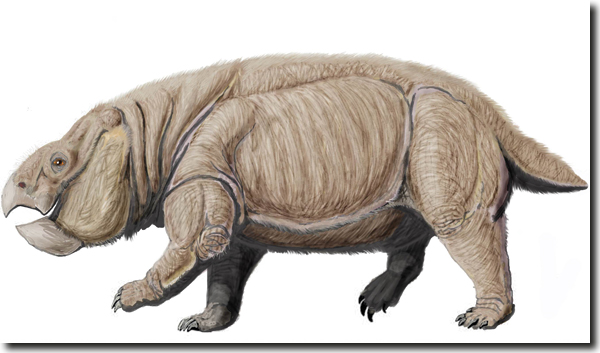
Abb. 5: Rekonstruktion von Lisowicia bojani, dem gigantischen Dicynodontier der späten Trias. (Bild: Dmitry Bogdanov, CC BY 3.0)
Anderseits wird nach Ansicht der Autoren klar, wie lückenhaft und unvollständig die hypothetische evolutionäre Geschichte durch Fossilien widergespiegelt wird. Lisowicia bojani erscheint geographisch und morphologisch unvermittelt mit einer nicht übermittelten Fundlücke von ca. 45 MrJ zu mutmaßlichen Vorfahren.
„The find of Lisowicia shows that at least one dicynodont lineage also participated in the ‘push for gigantism’ at the same time as the sauropodomorphs but also suggests that their evolutionary history in the Late Triassic is poorly documented …“ (Sulej et al. S. 364).
Konvergenzen
Lisowicia bojani kann als weiteres Beispiel für das innerhalb des Evolutionsparadigmas unverstandene, aber tausendfach belegte Faktum von Parallelentwicklungen und Konvergenzen angeführt werden. Wie bei den Dinosauriern und bei den Säugetieren finden sich bei Lisowicia bojani senkrecht unter dem Körper angeordnete Gliedmaßen (Säulengang). Das erfordert – wenn man ein evolutionäres Entstehungsszenario voraussetzt – eine gewaltige morphologische und physiologische Umgestaltung der Gliedmaßen im Vergleich mit seinen hypothetischen Vorfahren. Weiter muss postuliert werden, dieser Umbauprozess sei mindestens dreimal unabhängig voneinander bei den Dinosauriern, bei den Säugetieren und hier bei Lisowicia bojani im Laufe der Evolution „erfunden“ worden. Weitere Merkmale, die als Konvergenzen interpretiert werden müssen, sind das Phänomen des kontinuierlichen Größenwachstums wie bei Dinosauriern, das Vorhandensein von Aushöhlungen in Wirbelkörpern wie bei Sauropoden oder die sekundäre Umgestaltung des Gebisses in einen Schnabel aus Horn ähnlich wie bei den heutigen Schildkröten. Die bislang favorisierten und diskutierten Evolutionsmechanismen im Rahmen der Modellbildungen einer erweiterten evolutionären Synthese bzw. unter Zugriff auf moderne Ansätze von Evo-Devo können diese Befunde und Merkmalsverteilungen nicht einmal ansatzweise plausibel erklären.
Evolutionsmechanismen und Selektion
Nicht nur hinsichtlich der Erklärung von Konvergenzen, sondern auch unabhängig davon sind die o. g. Evolutionsmechanismen überfordert, die morphologischen und funktionellen Innovationen bei Lisowicia bojani zu erklären. Um riesig zu werden, musste diese Form der Dicynodontier gravierende Veränderungen in Körperbau und Stoffwechsel durchlaufen. Die Entstehung bzw. der Umbau von vorhandenen Bauplänen, sodass die Oberarmknochen senkrecht unter dem Körper stabil Platz fanden und funktionierten, erforderte eine andere Konfiguration der Muskulatur und damit innerhalb der Dicynodontier eine völlig neue Konstruktion. Parallel dazu bleibt offen, warum und wie in den Wirbelkörpern Aushöhlungen entstehen konnten oder wie rasches und dauerhaftes Knochenwachstum möglich wurde. Und das alles im Verlauf einer aus evolutionärer Perspektive vermeintlich relativ kurzen evolutionären Spanne von nur 25 Millionen Kalenderjahren, wie die Autoren vermuten (wobei radiometrische Jahre mit Kalenderjahren gleichgesetzt werden).
Die bekannten Evolutionsmechanismen sind überfordert, die morphologischen und funktionellen Innovationen bei Lisowicia zu erklären.
Nur am Rande werden von Sulej & Niedzwiedzki (2018) Aspekte der Selektion diskutiert, welche die Evolution von Lisowicia bojani vorangetrieben und kanalisiert haben könnten. Dass Lisowicia bojani sich zu einem riesigen Tier mit dem Körperbau eines Nashorns entwickelte, könnte an dem damit verbundenen Schutz vor Raubtieren (Dinosauriern) gelegen haben, spekulieren die Autoren. Vielleicht waren es auch Vorteile bei der Verwertung der Nahrung oder einer optimierten Verwertung von Energie, weil sich bei Riesenwachstum die Körperoberfläche, über die kostbare Energie in Form von Wärme verloren geht, im Verhältnis zur Körpermasse reduzierte. Allerdings muss immer wieder daran erinnert werden, dass ein Nutzen im fertig ausgeprägten Zustand keinerlei Erklärung für die Entstehungsweise liefert.
Abschluss
Fossilien offenbaren immer wieder eine erstaunliche und atemberaubende Formenvielfalt ausgestorbener Tiergruppen. Offenkundig ist die zunehmende Fülle von aus der Perspektive des Evolutionsparadigmas unerwarteten und nicht erklärbaren Daten wie beispielhaft bei Lisowicia bojani dokumentiert. Dazu zählen die extrem raschen Entwicklungen neuer Baupläne innerhalb sehr kurzer evolutionärer Zeitspannen, das Auftreten von tiefgreifenden Konvergenzen zwischen weit entfernten Tiergruppen, die Tatsache der fehlenden fossilen Dokumentation des vermuteten evolutionären Wandels und nicht zuletzt die unbeantworteten Fragen nach den Mechanismen und den Selektionsdrücken der Evolution. Diese Situation fordert eine grundsätzliche Infragestellung des Konzeptes der Evolution. Ein schöpfungstheoretischer Ansatz zur Deutung der Fossilen vermag manche der o.g. Probleme besser zu erklären, weil eine freie Kombinierbarkeit von Merkmalen und plötzlich auftretende Formen aus der Sicht von Schöpfung plausibel sind. Dieser Ansatz muss sich aber der nicht minder großen Herausforderung stellen, die geographische und zeitliche Ordnung bzw. Verteilung der Fossilien sowie die tiefgreifenden Wechsel der Ökosysteme, in denen z. B. die heute fremdartig anmutenden Dicynodontier und Dinosaurier zu Hause waren, besser zu deuten. Bis dahin ist es noch ein weiter Weg.
1 Amnioten sind Landwirbeltiere, deren Embryonen sich in einer mit Fruchtwasser (Amnionflüssigkeit) gefüllten Amnionhöhle entwickeln, die von einer Embryonalhülle (lat. Amnion) umgeben ist.
2 Dazu zählen z. B. Reptilien, Dinosaurier, Vögel und deren ausgestorbene Vertreter.
3 Tiere mit einem charakteristischen Knochenfenster an der hinteren Schädelseitenwand, zu denen die sehr umfangreiche Ordnung der Therapsiden gehört, welche die Säugetiere einschließt (vgl. Abb. 2).
An elephant-sized Late Triassic synapsid with erect limbs. Science 363, 78–80.
The last dicynodont: an Australian Cretaceous relict. Proc. R. Soc. Biol. Sci. 270B, 985–993.
Evolution – ein kritisches Lehrbuch. Gießen, 7. Aufl.
Themen | Kurzbeiträge | Streiflichter
Studiengemeinschaft WORT und WISSEN e.V.
Letzte Änderung: 6/25/21
Webmaster