
Kretazische Insekten im Gestein und als Einschlüsse
Stratigraphische Besonderheiten weisen auf kurze erdgeschichtliche Zeiträume hin
von Michael Suling
Studium Integrale Journal
23. Jahrgang / Heft 1 - Mai 2016
Seite 13 - 22
Zusammenfassung: Die zeitliche Verteilung von Insektenfossilien im geologischen System der Kreide und die Art ihrer Konservierung (zusammengepresst und mineralisisert oder in Bernstein) zeigt auffällige Muster. Die fossile Überlieferung in Bernstein und als Kompressionsfossil ist markant unterschiedlich. Diese Unterschiede sind im Rahmen kurzer Zeiträume besser zu verstehen als in einem Langzeitkonzept.
Ein fossiler Netzflügler (Neuroptera: Araripeneuridae) aus der kretazischen Crato-Formation, Brasilien. (© Dr. James E. Jepson, mit freundlicher Genehemigung des Autors und von Siri Scientific Press; aus Penney & Jepson 2014) Einleitung
Die Insekten stellen eine Klasse innerhalb der Hexapoda (Sechsfüßer) dar. In diesem Unterstamm der Arthropoda (Gliederfüßer) werden als weitere Klassen die Diplura (Doppelschwänze), die Collembola (Springschwänze) und die Protura (Beintaster) unterschieden.
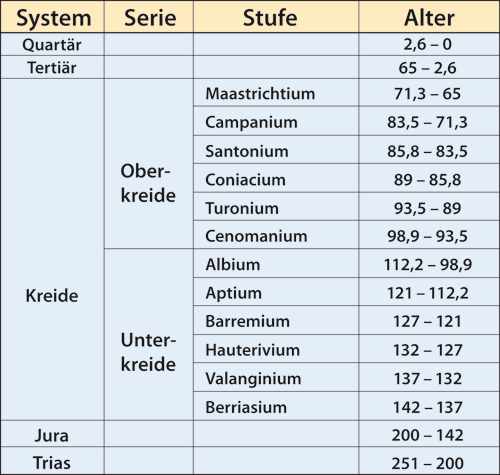
Abb. 1: Unterteilungen des Systems Kreide. Angaben zum Alter in Millionen radiometrischen Jahren.
Exemplare von fossilen Insekten liegen heute in einer nicht mehr überschaubaren Anzahl vor. In den letzten vier Jahrzehnten hat die Erschließung vieler neuer Fundstellen inbesondere in kretazischen* Sedimenten zu einer wesentlich besseren Kenntnis der damaligen Insektenwelt geführt. So wurden allein aus der Fundstätte Baissa Creek in Sibirien, die heute ins Aptium (vgl. Abb. 1) gestellt wird, bis zum Jahr 2000 ca. 20 000 Exemplare von Insekten geborgen. Weitere große Deponien fossilisierter Lebewesen etwa gleichen Alters liegen bei Bon Tsagan in der Mongolei und im brasilianischen Araripebecken. Paläontologen konnten hier jeweils über 10 000 fossile Insekten sammeln. Tausende Fundstücke sind noch unbeschrieben und beinhalten vermutlich noch manche Überraschung.
Für statistische Aussagen über das Vorkommen fossiler Insekten bietet sich als taxonomische Ebene die Familie an. Insektenfamilien weisen oft eine große stratigraphische* Reichweite auf. So existieren ca. 84 % der aus dem Albium (vgl. Abb. 1) bekannten Familien auch heute noch. Insgesamt weist der Fossilbericht der Insekten gegenwärtig über 1100 Familien auf. Diese Anzahl hatte sich in der Zeit von 1983 bis 2005 um etwa 500 erhöht (Grimaldi & Engels 2005, 2). Etwa 68 % der ca. 800 heute lebenden Insektenfamilien sind auch fossil bekannt. Auf der Gattungsebene liegt dieser Anteil dagegen unter 2 %. Bei fossilen Exemplaren, die nicht im Bernstein vorliegen, ist eine sichere Bestimmung der Gattung in der Regel kaum möglich.
Obwohl Fossilien von Insekten in mehreren verschiedenen Formen auftreten, werden in diesem Beitrag nur zwei Einbettungsweisen unterschieden: Insekteneinschlüsse im Bernstein (engl. amber) werden als A-Fossilien (bzw. A-Dokumente) bezeichnet; alle anderen Einschlüsse fallen unter den Sammelbegriff C-Fossil (compression fossil; bzw. C-Dokument). Bei den C-Fossilien handelt es sich in der Regel um lakustrine* oder seltener um fluviatile* Einbettungen bzw. Ablagerungen. Die Muster der Verteilung der kretazischen A- und C-Dokumente sind überraschenderweise deutlich unterschiedlich. Dieser Befund ist Gegenstand der folgenden Untersuchung.
Im Fossilbericht der Insekten heben sich plötzlich auftretende größere Auslöschungen (mutmaßliches Aussterben) von Familien deutlich ab von einem relativ geringen sogenannten Hintergrund-Aussterben, dessen Rate im Mesozoikum gleichbleibend bei etwa 10 Familien pro Stufe liegt (Labandeira 2005). Diese Situation erweist sich für Deutungen statistischer Merkmale bei Vorkommen von fossilen Insekten als vorteilhaft.
Datenmaterial
Den folgenden Betrachtungen liegen im Wesentlichen Informationen aus der EDNA FOSSIL DATA BASE und der PALEOBIOLOGY DATABASE (PBDB) zugrunde. Beide Quellen geben den Bestand an bekannten fossilen Exemplaren allerdings nur unvollständig wieder. Daher wurden darüber hinaus weitere Fundmitteilungen aus zahllosen Veröffentlichungen (z. B. Penney 2010, Perrichot et al. 2007, Grimaldi et al. 2002, Makarkin et al. 2012) berücksichtigt. Natürlich ist eine annähernd lückenlose Erfassung von Insektenfossilien nicht möglich. Innerhalb der meisten Insektenordnungen sind weit mehr Exemplare bekannt, als die Anzahl der beschriebenen Arten vermuten lässt. Man darf aber angesichts der in diesem Beitrag verwendeten Datenfülle die darin auftretenden Relationen zwischen den Anzahlen der Taxa in der Regel als hinreichend signifikant für die fossile Überlieferung betrachten.
Die taxonomischen Bestimmungen der einzelnen Exemplare und ihre Einordnung in die geologische Zeitskala orientieren sich an den Zuordnungen der PBDB.
Kompakt
Unterzieht man die zeitliche Verteilung sowie die Konservierungsmodi von fossilen Insekten einer einfachen statistischen Betrachtungsweise, so stößt man auf mehrere Auffälligkeiten:
• Bei Insekten findet – auf der Ebene der Familie – nach Abschluss des Perms nur noch am Ende des Aptiums (Stufe der Unterkreide, vgl. Abb. 1) eine wesentliche Auslöschung statt. Während diese bei sedimentären* Einschlüssen sehr markant hervortritt, wird sie merkwürdigerweise von den Inklusen* des Bernsteins kaum widergespiegelt.
• Sehr viele der heute noch lebenden Familien, die man vor dem Albium (ebenfalls Stufe der Unterkreide, Abb. 1) allein aus Sedimenteinschlüssen kennt, sind in der Oberkreide gar nicht oder allein als Bernsteineinschlüsse nachgewiesen.
• Während die Familienvielfalt bei sedimentären Einbettungen an der Aptium-Albium-Grenze einen deutlichen Rückgang wenigstens bis zum Beginn des Tertiärs erkennen lässt, zeichnet sich bei den Bernsteininklusen in der mittleren Phase der kretazischen* Sedimentation eine starke Zunahme ab.
Diese gegenläufigen Tendenzen in der fossilen Überlieferung von Insektenfamilien im Bernstein einerseits und in Sedimenten andererseits führen zu einer Hinterfragung der für die oberkretazischen Bernsteininklusen angegebenen Ablagerungsbedingungen und Entstehungszeiten. In einem kurzen Zeitrahmen sind Deutungen der Fundsituation fossiler Insekten teilweise weniger problematisch.

Abb. 2: Der Rüsselkäfer Burmonyx zigrasi (Nemonychidae; ca. 2,3 mm groß; Bernstein aus Myanmar). Die rezente Familie Nemonychidae ist auch aus dem libanesischen Bernstein (Barremium; s. Abb. 1) sowie aus dem Jura von Australien, Kasachstan und der Mongolei bekannt. (http://sciencythoughts.blogspot.de/2014/05/a-pine-cone-weevil-from-cretaceous.html) Räumliche und zeitliche Unstetigkeiten

Abb. 3: Eine Libelle der auch heute vorkommenden Familie Thaumatoneuridae. Diese Familie findet sich häufig in der Crato-Formation (Aptium). Dokumente aus dem Bereich der C-Lücke sind nicht bekannt. (Aus Bechly 2010)
Weltweit sind mehrere hundert Fundstellen fossiler Insekten bekannt, die sich auf nahezu alle Stufen oberhalb des Devons verteilen. Auffällig ist aber, dass diese Fossilien in nur wenigen Lagerstätten besonders häufig auftreten, während die meisten Fundstellen nur wenige Fossilien bergen. Einige Beispiele mögen dies verdeutlichen:
• Im Campanium (Oberkreide; Abb. 1) sind Insekten praktisch nur aus dem kanadischen Bernstein – hier mit über 110 verschiedenen Familien – und von der russischen Ola-Formation bekannt.
• Insekten des Santoniums, der nächst älteren Stufe, finden sich fast ausschließlich in Sibirien und liegen hier ebenfalls als Bernsteineinschlüsse vor.
• Im Bereich kretazischer Ablagerungen ist in Brasilien außer den überreichen Funden im Araripebecken (Aptium/Albium) nur noch eine geringe Anzahl an Insektenfossilien bekannt. Ähnliches gilt für die entsprechenden kretazischen Nachweise in Australien, die sich auf die ins Aptium gestellte Koonwarra-Formation konzentrieren.
Nach Labandeira & Eble (2000, 6) ist diese episodenartige Form erdgeschichtlicher Überlieferung fossiler Insekten wesentlich bestimmt durch die jeweiligen Lagerstätten und zeigt sich in ausgezeichnet erhaltenen reichen Ablagerungen, die unregelmäßig in mancherlei grundverschiedenen Umfeldern auftreten.
Solche ausgeprägten räumlichen und zeitlichen Unstetigkeiten stellen kritische Anfragen an die Vorstellung von Insekteneinbettungen im Rahmen einer nur sehr langsam fortschreitenden Sedimentation dar. Die vorliegende Form dieser Überlieferung kann auch und vielleicht besser im Kontext von katastrophischen Ereignissen verstanden werden.
Die Fossilüberlieferung bezüglich der Familien
Zeitliche Verteilung von fossilen Insekten
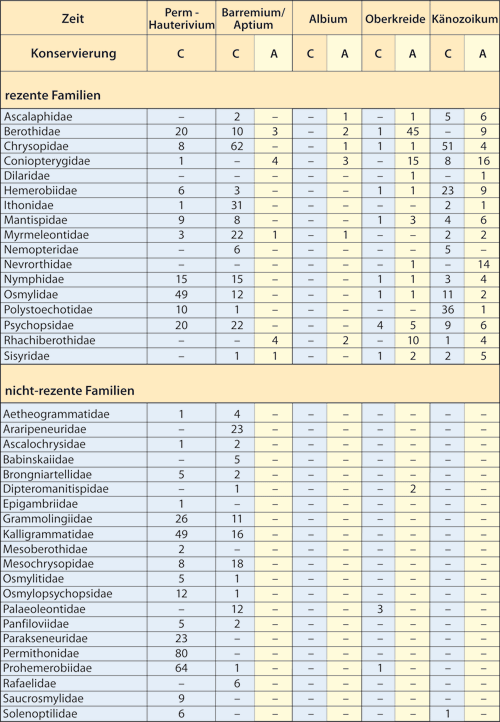
Abb. 4: Häufigkeiten der Fossilien der Familien der Ordnung Neuroptera in verschiedenen Phasen der Erdgeschichte. A: Harz-Einbettung, C: harzfreie Einbettung. Unerwartet sind die auffälligen Unterschiede in den Häufigkeitsverteilungen rezenter und nicht-rezenter Familien.
Aus Abb. 4 ist ersichtlich, mit welcher Häufigkeit Familien der Ordnung Neuroptera in verschiedenen Phasen der Erdgeschichte auftreten. Natürlich ist bei allen Graphiken bezüglich des Vorkommens fossiler Taxa zu beachten, dass Lebensdauer und -raum dieser Taxa oft weit ausgedehnter gewesen sein dürften, als es die gefundenen Nachweise anzeigen. Dennoch darf man in den sich deutlich abzeichnenden Verteilungsmustern aufgrund der großen Datenfülle mehr als nur zufällige Erscheinungen sehen.
In Abb. 4 fallen bezüglich der Neuroptera mehrere Besonderheiten im stratigraphischen Bericht auf, z. B.:
Diese Verteilung, die sich auch innerhalb anderer Insektenordnungen abzeichnet, ist keineswegs zu erwarten:
Nachfolgend soll nun zunächst ein genaueres Bild der Dokumentation kretazischer Insekten gegeben werden, bevor die auffälligen Muster der Fossilüberlieferung kritisch diskutiert werden.
C-Fossilien und A-Fossilien
Etwa 35 % der ca. 720 Insektenfamilien, die aus kretazischen Ablagerungen bekannt sind, liegen hier sowohl als C- als auch als A-Dokumente vor. Die bei fossilen Insekten erkennbaren stratigraphischen Besonderheiten treten nur dann deutlich hervor, wenn man nicht allgemein nach den stratigraphischen Vorkommen von Familien fragt, sondern die Verteilung der C- und A- Komponenten ihrer Fossilien jeweils gesondert betrachtet.
C-Fossilien
Am Ende des Aptiums ist ein sehr markanter Rückgang der Anzahl der C-Fossilien erkennbar. Vorausgegangen war eine deutliche Zunahme seit Beginn der Kreide. Berücksichtigt man allein die in der PBDB gemachten Angaben, so stehen den 3016 C-Exemplaren des Aptiums nur 27 (!) im Albium gegenüber. Betroffen von diesem Einschnitt in der C-Dokumentation sind – fast ausnahmslos – alle Familien, von denen im Aptium C-Dokumente vorliegen. Innerhalb der etwa 396 Familien, die in dieser Stufe – neben eventuellen Einschlüssen im Bernstein – durch C-Fossilien vertreten sind, lassen sich drei paarweise disjunkte Gruppen unterscheiden:
Es fällt auf, dass praktisch alle 150 Familien, die im Barremium bzw. Aptium als C-Dokument vorliegen und oberhalb der Abbruchkante nicht mehr existieren, in dem diesen Stufen zugeordneten libanesischen Bernstein fehlen – ein sonderbarer Befund (vgl. dazu nachfolgenden Abschnitt „A-Fossilien“ und Abschnitt „Weitere Aspekte“).
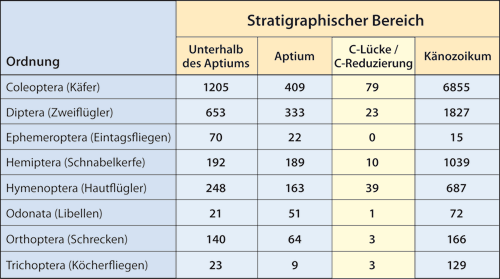
Abb. 5: Summarische Häufigkeiten von C-Dokumenten für Familien innerhalb verschiedener Ordnungen (Stand: Januar 2016). Berücksichtigt sind nur Familien, die im Aptium als C-Fossil auftreten und noch heute existieren. Die Daten orientieren sich an den Angaben der PBDB.
Innerhalb dieser dritten Gruppe gibt es ca. 75 Familien, die die C-Lücke überleben. Praktisch alle diese Familien liegen als C-Exemplare im Aptium nie seltener, aber oft sehr viel häufiger als in der breiten C-Lücke vor (Abb. Z3). Damit zeichnet sich bei diesen Familien parallel zur C-Lücke eine markante C-Reduzierung ab.
Der durch die C-Lücke und die C-Reduzierung bedingte Einschnitt in der C-Komponente ist für einige Insektengruppen aus Abb. 5 ersichtlich.
Die C-Lücke wird vergrößert durch weitere ca. 50 rezente Familien, bei denen die bis ins Känozoikum reichende Unterbrechung der C-Dokumentation schon vor dem Aptium einsetzt.
A-Fossilien
Bei den A-Fossilien ist mit Beginn des Barremiums ein sprungartiger Anstieg sowohl der Familien-Vielfalt als auch der Anzahl der Exemplare festzustellen. Im weiteren Verlauf der Kreide kommt eine Fülle von Inklusen hinzu.
Von den im Bereich der C-Lücke insgesamt fossil nachgewiesenen (nicht gleichbedeutend mit „existierenden“) etwa 430 Familien kommen ca. 90 % in Bernsteineinschlüssen vor und nur ca. 25 % als C-Fossilien. Bezogen auf die Anzahl der Exemplare ist der Anteil der C-Fossilien hier noch wesentlich geringer.
Hinweis: Die Abb. Z-Verweise beziehen sich auf zusätzliche Abbildungen unter insekten.pdf.
Diskussion
Rahmenbedingungen
Zunächst soll ein Rahmen beschrieben werden, in den sich Deutungen der komplexen Fundsituation einordnen lassen müssen:
Ein naheliegendes Szenario
Insbesondere eventstratigraphische* Untersuchungen legen nahe, dass im Verlauf der Kreide der Meeresspiegel enorm angestiegen ist (Haq 2014). So wird davon ausgegangen, dass in der Oberkreide ein Drittel der damaligen Landflächen unter Wasser gerieten. Das Maximum der Überflutung wurde aber nicht auf allen Kontinenten gleichzeitig erreicht. Als breiter Meeresarm durchzog der Western Interior Seaway den nordamerikanischen Kontinent von Norden nach Süden (Blakey 2014). In diesem ausgedehnten epikontinentalen Flachmeer, das während der gesamten Oberen Kreide bestand, sind heute viele marine Wirbeltiere eingebettet. Auf dem amerikanischen Südkontinent griff das Weltmeer von Norden her tief in das Landesinnere ein und hinterließ mehrere hundert Meter mächtige kreidereiche Ablagerungen, die u. a. viele große Sauropoden bergen. Über weite Gebiete im Westen Afrikas erstreckte sich ein Meer, das die Tethys (ein im Bereich des heutigen Mittelmeeres gelegener Ozean) mit dem südlichen Atlantik verband. Insgesamt erfolgte weltweit eine Verschiebung von terrestrischer* zu mariner Sedimentation.
Die Oberkreide ist durch weitere katastrophische Ereignisse gekennzeichnet. Es herrschte ein ungewöhnlich heftiger Vulkanismus, der teilweise mit der Plattenverschiebung verbunden war. Auf dem Festland und auf den Ozeanböden entstanden große Massen basaltischer Gesteine. Im Cenomanium schlug in Nordamerika am Rande des Western Interior Seaway ein Metorit ein, dessen Durchmesser auf 2 km (!) geschätzt wird. Der hinterlassene Krater liegt heute unter der Stadt Manson (Iowa) und hat eine Breite von ca. 38 km (Koeberl & Anderson 1996).
Man darf annehmen, dass die skizzierten geologischen Ereignisse zur Vernichtung vieler Lebensräume und in deren Folge zu massiven Dezimierungen innerhalb der Insektenpopulationen und zum Aussterben vieler Familien führten. Mit fortschreitender Transgression des Meeres wurden die Möglichkeiten einer lakustrinen* Einbettung wesentlich eingeschränkt. Durch diese Ereignisse wird zunächst der extreme Rückgang der C-Fossilien mit dem Beginn des Albiums verständlich. Vermutlich spiegelt der vorausgehende Boom an C-Fossilien ein Massenbegräbnis wider, das zur Entstehung der Abbruchkante, der C-Lücke sowie der C-Reduzierung beitrug.
Solche Szenarien lassen sich im Rahmen kurzzeitiger katastrophischer Vorgänge gut verstehen. Dagegen lässt sich eine gemäß Abb. 1 bestehende Dauer von 50 Ma, während derer die C-Dokumentation unterbrochen oder erheblich reduziert ist, nicht überzeugend erklären, weder durch Dezimierungsprozesse noch durch Einschränkung lakustriner Sedimentation:
Die Unterbrechung beziehungsweise die erhebliche Reduzierung der C-Dokumentation lässt sich für eine Dauer von 50 Ma nicht überzeugend erklären.
Insbesondere ist es kaum denkbar, dass die Regeneration der Insektenpopulationen erst 50 Millionen Jahre nach der etwa mit dem Ende des Aptium einsetzenden mächtigen Ausbreitung der zuvor nur spärlich vorhandenen Angiospermen (Bedecktsamer) einsetzte. Denn einerseits wurde mit dieser Entwicklung der Flora das Nahrungsangebot für Insekten erheblich erweitert. Von den heutigen Insektenarten sind ca. 43 % phytophag* (Grimaldi & Engels 2005, 622). Andererseits sind auch viele Angiospermen für ihren Fortbestand auf Insekten angewiesen. Nach Grimaldi & Engels (2005, 5) werden annähernd 85 % der heutigen ca. 250 000 Arten der Angiospermen durch Insekten bestäubt.
Bei Annahme sehr langer Zeiträume bestanden offensichtlich viele Biotope und Zeitphasen, in denen zwischenzeitliche Regenerationen vielfältig hätten erfolgen können, und es existierten auch genügend Räume für sedimentäre Einbettungen. Das auffällige Fehlen von C-Fossilien wirft daher die Frage auf, welche reale Zeitdauer der C-Lücke aufgrund der genannten Befunde zugestanden werden kann.
Die Dominanz der A-Fossilien
Im aktualistischen Langzeit-Konzept der Erdgeschichte erweist sich aber nicht nur die Deutung der C-Lücke sowie der C-Reduzierung als problematisch. Rätselhaft bleiben weitere Befunde:

Abb. 6: Die Raubfliege Burmapogon bruckschi (Familie Asilidae) aus burmesischem Bernstein. Die rezente Familie Asilidae ist auch aus dem Aptium von Brasilien und Sibirien sowie dem Jura von Kasachstan bekannt. Sie erscheint in der C-Lücke nur als A-Fossil. (http://www.sci-news.com/paleontology/science-assassin-flies-burmese-amber-01874.html, © David Grimaldi) Die sehr zahlreichen Familien des libanesischen Bernsteins lassen durchaus kein Aussterbeereignis am Ende des Aptiums erkennen. Hier besteht ein markanter Gegensatz zu dem plötzlichen endgültigen Verschwinden vieler Familien aus der alleinigen C-Dokumentation des Aptiums. Dadurch zeichnet sich an der Grenze zum Albium eine recht paradoxe Situation bezüglich des Überlebens ab.
Der während der C-Lücke bestehende starke Kontrast zwischen den in großer Anzahl und Vielfalt vorliegenden Inklusen und den vergleichsweise wenigen C-Dokumenten führt zur Frage nach der Herkunft der Harzeinschlüsse. Die mit dem Ende des Aptiums einsetzende Divergenz zwischen den beiden Konservierungsweisen verliert ihre Problematik, wenn man die Inklusen als Verdriftungen aus Biotopen ansieht, die schon in der Zeit vor dem Albium bestanden. Delclos et al. (2007) räumen ein, dass die Harzlagerstätten kaum als autochthon* angesehen werden können: „… gewöhnlich wird der Bernstein nicht am Ort seiner Entstehung gefunden, sondern nach Umlagerung durch Wasserströme zu den endgültigen Lagerstätten der Sedimente.“
Auch diese Schlussfolgerung, dass Bernsteine durch Transport verdriftet werden, ist im Rahmen einer kurz währenden, katastrophisch verlaufenden Erdgeschichte gut verstehbar.
Fossile Harze werden häufig im Umfeld von Kohlevorkommen gefunden. Die Ursache hierfür kann u. a. darin gesehen werden, dass Harze und Vegetationsmassen als schwimmfähige Materialien ähnliche Transporteigenschaften aufweisen und so gleichzeitig als Strandgut hinterlassen werden können. Diese Vorstellung wird – trotz mancher Verweise auf autochthone* Bernsteinvorkommen in Kohlen – nicht grundsätzlich bestritten: „Die weltweit wesentlichen Bernsteinlagerstätten treten in Form von Delta-Ablagerungen auf, die in der Regel reich an kohlehaltigem Schluffstein sind ... Durch Flüsse transportierte Harze wurden zusammen mit pflanzlichen Überresten von Wäldern in Deltaebenen angehäuft“ (Gomez et al. 2002). Eine Allochthonie* des Bernsteins lässt zunächst die Autochthonie der diese Harze beherbergenden Kohlen als fraglich erscheinen, dann aber auch die Autochthonie der Sedimente, in denen dieses fossile Material gefunden wird.
Die sich anbietende Annahme, dass es sich bei den Inklusen der C-Lücke um verdriftetes Material aus früheren Biotopen handelt, ist wenigstens verträglich mit Beobachtungen von Umlagerungen bei kretazischen Harzen. Darüber hinaus stellt diese Annahme aber auch erneut die langen Ablagerungszeiten in Frage, die den kretazischen Sedimenten üblicherweise zugeordnet werden.
Einige Beispiele möglicher Einschwemmung
Die Lagerungsverhältnisse einiger bedeutender Harz-Fundstellen lassen eine Entstehung durch Einschwemmung von Inklusen erkennen oder vermuten.
Kanada: Die Allochthonie der Bernsteine der bedeutenden kanadischen Lagerstätte Cedar Lake (dem Cenomanium zugeordnet) wird heute kaum bezweifelt. Diese Bernsteine gelten als ein über viele hundert Kilometer umgelagerter Teil der fossilen Harze von Grassy Lake in SE-Alberta (McKellar et al. 2008).
Über die Entstehung der Grassy Lake-Inklusen selbst schreiben McKellar et al. (2013): „Der kanadische Bernstein entstand etwa vor 78-79 Millionen Jahren, als zypresssenähnliche Bäume, die an den Rändern des Western Interior Seaways lebten, Harze freisetzten, die von den Ablagerungen naher Salzmarschen oder Küstenseen aufgefangen wurden. Diese Ablagerungen verfestigten sich und wurden schließlich ein Teil des Bereichs der Taber-Kohle ... .“
Der zitierte Harzlieferant aus der Familie der Cupressaceae ist jedoch gemäß PBDB im Bereich von Grassy Lake praktisch nicht fossil vorhanden und es fehlt der Nachweis der Autochthonie des Bernsteins. Gerade die exponierte Lage „along the margins of the ... Seaway“ kann als Hinweis dafür angesehen werden, dass die Pflanzenmassen der Tabor-Kohle auf einander folgenden Horizonten gestrandet sind und der Bernstein als ebenfalls schwimmfähiges Material einfach ein ähnliches Schicksal erfuhr. Die fossilen Harze überlagern hier ein 70 cm mächtiges Kohleflöz.
Insgesamt gehören zur Tabor-Kohle sechs Kohleflöze. Mit Verweis auf Pike (1995) wird von McKellar et al. (2008) eine autochthone Entstehung dieser Flöze hinterfragt: „... diese Kohlenflöze stellen wahrscheinlich lokale Abfolgen von Ablagerungen in Küstenseen und Salzmarschen dar ... .“
Spanien: Auch der spanische Bernstein befindet sich teilweise in der Nähe von Kohlelagern. Die Vorkommen der dem Albium zugerechneten harzführenden Aufschlüsse (Abb. Z6) lokalisieren Delclos et al. (2007) wie folgt: „Diese Aufschlüsse verteilen sich auf einen gebogenen Streifen, der vom Norden zum Osten der Iberischen Halbinsel verläuft und der die Küstenlinie während der Unterkreide darstellt.“
Die Positionierung dieser Bernsteine weit entlang einer kreidezeitlichen Küste unterstützt die Vermutung einer allochthonen Entstehung dieser Lagerstätten. Kaum einen Zweifel an dieser Sichtweise lassen insbesondere die bernsteinführenden Ablagerungen bei El Soplao (Basque-Cantabrian-Basin), deren Entstehung Najarro et. al. (2009) so beschreiben: „Die Las Penosas-Formation gehört zur regressiven Phase dieser Schichtenfolge … Die Schichten, die Kohle und Bernstein führen, kommen im Bereich von Deltamündungen vor, die sich während des Maximums der regressiven Abläufe bildeten …“ Und nach einem Verweis auf die Ähnlichkeit aller unterkretazischen Bernsteinlager in Spanien konkretisieren die Autoren die Fundsituation wie folgt: „... das Umfeld der Ablagerungen von El Soplao weist einen leichten Meereseinfluss auf, wie die Vorkommen einiger kleiner mariner Muscheln und Schnecken zwischen den Bernsteinablagerungen zeigen und wie auch Bryozoen und Serpuliden, die Krusten an der Oberfläche einiger Bernsteine bilden, erkennen lassen.“
Jordanien: Die Bernsteine der im Wadi Zerqa aufgeschlossenen harzführenden Gesteine (heute dem Albium zugeordnet) sind zusammen mit Vegetationsresten (u.?a. Blätter von Angiospermen) den hier etwa 320 m (!) mächtigen Sedimenten des Kurnub-Sandsteins eingelagert, deren Material von Süden her antransportiert wurde (Bandel & Salameh 2013).
„Es handelt sich hier um Ablagerungen eines Küstenwaldes am Randsaum des Arabisch-Afrikanischen Kontinents, dessen Waldboden durch wiederholte Meeresvorstöße aufgearbeitet wurde, wobei die herausgewaschenen Harze nach mehr oder weniger langem Aufenthalt im marinen Milieu nahe ihres Entstehungsortes wieder im Sediment abgelagert wurden“ (Krumbiegel & Krumbiegel 2005, 58).
Wann und wo bestanden harzreiche Biotope?
Eine Übersicht über die Verbreitung von Aufschlüssen, in denen die postulierten Harzlieferanten Araucariaceae, Taxodiaceae, Cupressaceae, Cheirolepidiaceae nachgewiesen wurden, ist in Abb. Z5a-d wiedergegeben. Zugrunde gelegt wurde hierbei das Datenmaterial der PBDB. Diese vielen Fundstellen führen fast immer nur äußerst wenige häckselförmige Reste der genannten Pflanzen und vermitteln schwerlich den Eindruck, dass man hier einen Einblick in eine autochthone Vegetation hat. Regionen, die eine weiträumigere Anhäufung solcher „mageren“ Aufschlüsse aufweisen, stehen in der Regel in großer räumlicher und zeitlicher Distanz zu den wesentlichen Bernsteinlagerstätten. Die genannten Distanzen treten noch stärker hervor, wenn man nach einer Beziehung der einzelnen Lagerstätte (Abb. Z5a-d) zu der als Quelle vermuteten speziellen Pflanzenfamilie sucht. Auch wenn für mehrere fossile Harze die pflanzlichen Lieferanten bis heute nicht sicher bestimmt werden konnten, so stützt doch das Fehlen einer signifikanten Korrelation von Harzlagern und Harzproduzenten die Sichtweise einer allochthonen Entstehung dieser Lagerstätten.
Gemäß Abb. Z5a-d finden sich Nachweise fossiler Harzlieferanten in oberkretazischen Sedimenten nicht zahlreicher als in oberjurassischen. Aus den zuletzt genannten Ablagerungen sind aber nur wenige Bernsteinfunde bekannt und Inklusen fehlen fast vollständig. Offen bleibt die Frage, ob hier eine Deponierung fossiler Harze kaum stattfand, ob abgelagertes Material zerfiel oder ob es später umgelagert wurde.
Weitere Aspekte
1. Es stellt sich natürlich die Frage, weshalb am Ende des Aptiums Dezimierungen und Auslöschungen im Wesentlichen nur bei den Hexapoda und Spinnentieren feststellbar sind und nicht auch etwa bei Landwirbeltieren. Nun waren die Sauropoden, die gemäß den fossilen Zeugnissen die terrestrische Fauna der Unterkreide dominierten, in der Regel an ein Leben im Bereich von interkontinentalen Flachmeeren gut angepasst. Plazentale Säugetiere sind aus dieser Phase der Erdgeschichte allgemein kaum nachgewiesen.
2. Unter den C-Dokumenten des Barremiums und Aptiums sind Vertreter der Heuschrecken (Orthoptera), Libellen (Odonata), Eintagsfliegen (Ephemeroptera), Skorpionsfliegen (Mecoptera), Steinfliegen (Plecoptera), Köcherfliegen (Trichoptera), Wasserwanzen (Nepomorpha), Wasserläufer (Gerromorpha) sowie der Adephaga (im Wasserbereich oder am Boden lebende Käfer) zahlreich zu finden. Dagegen fehlen die Familien dieser Gruppe, die aus Bewohnern von eher tiefer gelegenen Ebenen und Feuchtbiotopen besteht, unter den zeitgleich abgelagerten Inklusen des Libanon fast vollständig. Dieses Fehlen ist auch deshalb bemerkenswert, weil innerhalb der Fauna des baltischen Bernsteins aquatisch lebende Insekten ca. 25 % der Inklusen stellen (Wichard et al. 2009, 17). Die Verschiedenartigkeit der beiden frühkretazischen Faunen ist sicherlich vorwiegend durch unterschiedliche Lebensräume bedingt. Azar (2007) stellt fest: „... die im libanesischen Bernstein eingeschlossene Fauna besteht größtenteils aus Tieren, die im unteren sowie mittleren Bereich von Bäumen lebten.“
Die extreme Dominanz der A-Dokumente in der C-Lücke kann durch Einschwemmung aus älteren Ablagerungen gedeutet werden.
Als recht sonderbar erscheint nun die Tatsache, dass die Familien der ersten Gruppe – im markanten Gegensatz zu jenen, die im libanesischen Bernstein vorkommen – am Ende des Aptiums stark von Auslöschungen betroffen wurden (s.o.). Das könnte seine Ursache darin haben, dass die zugehörigen Familien der Transgression des Meeres leicht zum Opfer fielen. Hingegen ist es vorstellbar, dass die Familien des libanesischen Bernsteins in Biotopen überlebten, die hinreichend hoch lagen und teilweise von harzreichen Bäumen besiedelt wurden. Jedenfalls gedeihen heutige Vertreter kretazischer Koniferenfamilien gut in Höhenlagen. Das Fehlen einer Abbruchkante, einer Lücke und einer Reduzierung bei den Familien dieser Inklusen würde dann verständlich. Aus dieser Vermutung lässt sich aber nicht folgern, dass den im Barremium bzw. im Aptium gefundenen Inklusen nicht eine schon frühere Entstehung zukommt. Die von Azar (2007) gegebene Beschreibung der Bernstein-Aufschlüsse deutet an, dass wenigstens ein Teil dieser fossilen Harze als allochthon betrachtet werden muss: „Der Bernstein des Libanons tritt ... gelegentlich in einem System reiner Flussablagerungen auf, d. h. in Rinnen oder an Ufern, und manchmal zeigt die Ablagerung marine Einflüsse ... Bei Flussläufen ... kann es vor allem nach Stürmen zur Anhäufung von Bernstein kommen, ... dagegen werden marine Schnecken bei Übergriffen des Meeres ins Sediment eingebracht.“
3. Auf das Verschwinden zahlreicher Familien am Ende des Aptiums nehmen Rasnitsyn & Quicke (2002, 416) Bezug: „... fast alle heute ausgestorbenen aquatisch lebenden Familien verschwanden gegen Ende der Unterkreide ... Nach einer vorläufigen Erklärung für dieses Auslöschungsereignis um die Mitte der Kreide ... wird eine Eutrophierung angenommen, die vom Land her erfolgte aufgrund der Ausbreitung der Angiospermen, die große Mengen schnell zerfallender Streu erzeugten.“
Die genannte Hypothese (Eutrophierung*) bezüglich der Ursache dieses Massen-Aussterbens vermag aber nicht zu überzeugen:
4. Aus der Annahme, dass die in der C-Lücke auftretenden Inklusen schon vor der Absetzung albischer Sedimente gebildet wurden, lässt sich eine testbare Folgerung ableiten: Außer den in der C-Lücke oft zahlreich vorkommenden A-Komponenten derjenigen Familien, die nach aussetzender C-Dokumentation während der C-Lücke dann wieder im Känozoikum nachgewiesen sind, sollte es Familien geben, deren C-Komponente (spätestens) an der Abbruchkante - durch angenommenes Aussterben der Familie - endgültig erlischt, die aber allein während der C-Lücke noch in Form eines „A-Überhanges“ auftreten. Tatsächlich weisen ca. 16 Familien (aus 8 verschiedenen Ordnungen) gerade diese Eigenschaft auf. Familien mit hierzu komplementären Eigenschaften bzgl. der A- und C-Dokumentation sind dagegen nicht bekannt.
Glossar
allochthon: nicht am Fundort entstanden. autochthon: am Fundort entstanden. Eutrophierung: Nährstoffanreicherung. Eventstratigraphie: Isochrone Gliederungsmöglichkeit durch plötzliche, in der Regel kurzfristige Ereignisse, welche Ausdruck in den Gesteinsablagerungen finden. fluviatil: auf Fließgewässer bezogen. Inkluse: Einschluss (hier: in Bernstein). Känozoikum: „Erdneuzeit“, beginnend mit dem Tertiär (vgl. Abb. 1). kretazisch: zum geologischen System der Kreide gehörend (vgl. Abb. 1). lakustrin: auf den Bereich eines Süßwassersees bezogen. marin: zum Meer gehörend. phytophage Insekten: Insekten, die sich von pflanzlichen Stoffen ernähren. rezent: heute lebend. sedimentär: in Ablagerungen befindlich. stratigraphisch: Die Abfolge der (geologischen) Schichten betreffend. terrestrisch: zur festen Landoberfläche gehörig.
Fazit
Etwa 160 auch aus dem Känozoikum bekannte Insektenfamilien, die im Aptium noch durch ihre C-Komponente überliefert sind, werden im Albium und in der Oberkreide in dieser Form nicht mehr gefunden. Weitere 127 Familien, die im Aptium als C-Dokument vorliegen, verschwinden am Ende dieser Stufe endgültig. Dieser extreme Einschnitt am Ende des Aptiums, der sich auch im Fehlen noch anderer C-Dokumente zeigt, bereitet im Rahmen großer Zeiträume, in denen der Ablauf geologischer Prozesse in der Regel als sehr langsam angenommen wird, erhebliche Verständnisschwierigkeiten. Weiter kontrastiert in der Oberkreide das Fortbestehen der Inklusen-Familien der frühen Kreide und die massive Zunahme der Familienvielfalt bei den Inklusen auffällig mit den Lücken bei sedimentärer Einbettung.
Durch dieses paradoxe Erscheinungsbild, aber auch durch die in der Regel von Verdriftungen gekennzeichneten Lagerungsverhältnisse des kretazischen Bernsteins und der ihn begleitenden Vegetation, müssen sowohl die Entstehungszeit dieser fossilen Harze als auch das Alter der sie bergenden Kohlen beziehungsweise Sedimente hinterfragt werden.
In einem Szenario, das die geologischen Ereignisse des Mesozoikums als kurzzeitige Katastrophen auffasst und das wenig Zeit für Regenerationsprozesse ließ, können die in der Unterkreide einsetzenden geologischen Veränderungen sinnvoll in einen ursächlichen Zusammenhang mit Dezimierungsprozessen gestellt werden, die dann auch teilweise zum Aussterben von Familien führten. In einem solchen Szenario lassen sich die kretazischen Bernsteine oberhalb des Aptiums zwanglos als aus älteren Biotopen stammende Einschwemmungen verstehen, die das Kreidemeer nach seiner Transgression zurückgelassen hat.
Die Fragen nach der Herkunft der fossilen Harze (Bernstein) müssen gesondert behandelt werden. Ebenso bleibt die Frage offen, weshalb die Familien der aptischen Inklusen fast ausschließlich zu solchen Familien gehören, die auch rezente Vertreter haben.
Dank
Dr. Reinhard Junker und Dr. Michael Kotulla danke ich für vielfältige Unterstützung.
Preservation and accumulation of biological inclusions in Lebanese amber and their significance. C. R. Palevol. 6, 151-156.
Geologic development of Jordan. Evolution of its rocks and life. Deposit No.690/3/2013 National library: 111-130. Amman.
Additions to the fossil dragonfly fauna from the Lower Cretaceous Crato Formation of Brazil (Insecta: Odonata). Paleodiversity 3, Suppl. 11-77.
Paleogeography and Paleotectonics of the Western Interior Seaway, Jurassic-Cretaceous of North America. Search and Discovery Article #30392.
Diversity of Hymenoptera and other insects in the Late Cretaceous (Turonian) deposits at Orapa, Botswana: a preliminary review. African Entomol. 11, 222.
Fossiliferous amber deposits from Cretaceous (Albian) of Spain. C. R. Palevol 6, 135-149.
Taphonomy and palaeoecology of plant remains from the oldest African Early Cretaceous amber locality. Lethaia 35, 300-308.
Fossiliferous Cretaceous amber from Myanmar (Burma): Its rediscovery, biotic diversity, and paleontological significance. Amer. Mus. Novit. 3361.
Evolution of the Insects. Cambridge University Press, New York.
Cretaceous eustasy revisited. Global and Planetary Change 113, 44-58.
The Manson Impact Structure, Iowa: Anatomy of an Impact Crater.Geological Society of America, Special Paper 302.
Bernstein. Fossile Harze aus aller Welt. Edition Goldschneck.
The Fossil Record of Insect Diversity and Disparity. SFI Working Paper: 2000-08-044: 6
The Fossil Record of Insect Extinction: New Approaches and Future Directions. American Entomologist 51, doi:10.1093/ae/51.1.14.
A comparative overview of the neuropteran assemblage of the Lower Cretaceous Yixian Formation (China), with description of a new genus of Psychopsidae (Insecta: Neuroptera). Cretaceous Res. 35, 57-68.
Correlation of Grassy Lake and Cedar Lake ambers using infrared spectroscopy, stable isotopes, and palaeoentomology. Can. J. Earth Sci. 45, 1061-1082.
New ants (Hymenoptera: Formicidae: Dolichoderinae) from Canadian Late Cretaceous amber. Bull. Geosci.88, 583–594.
Unusual concentration of Early-Albian arthropod-bearing amber in the Basque-Cantabrian Basin (El Saplao, Cantabria, Northern Spain): Palaeoenvironmental and palaeobiological implications. Geol. Acta 7, 363-387.
Biodiversity of fossils in amber from major world deposits. Siri Scientific Press, Manchester.
Fossil Insects: An introduction to palaeoentomology. Siri Scientific Press, Manchester.
A reassessment of the Cretaceous amber deposits from France and their palaeontological significance. Afr.Invertebr. 48, 213-227.
Amber taphonomy and the Grassy lake, Alberta, amber fauna. Ph.D. thesis. Department of Biological Science, The University of Calgary. Calgary, Alta.
Wasserinsekten im Baltischen Bernstein. Verlag Kessel.
History of insects. Kluwer Academic Publishers, Dordrecht.
Historische Geologie. Spektrum Akad. Verlag.