Wie sicher sind Deutungen in der Paläanthropologie? Das Beispiel Fußbeweglichkeit und zweibeinige Fortbewegung
Abb. 1: Das rechte Fußskelett des Menschen.
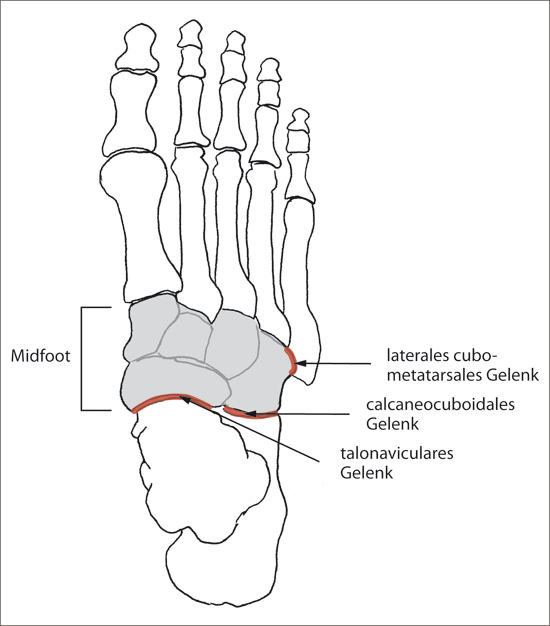
In der Diskussion um die vermutete Evolution des zweibeinigen menschlichen Ganges spielt die Beweglichkeit des Midfoot* (vorderer Fußwurzelbereich) eine große Rolle (Abb. 1).
Seit Jahrzehnten wird der sogenannte „midtarsale break“ – das ist eine Beugung des Midfoot nach plantar (zur Fußsohle hin) – als wesentliches Merkmal nichtmenschlicher Primaten bei zweibeiniger Fortbewegung angesehen. Dieser „Bruch“ wird typischerweise während der Standphase des Gehens beschrieben, wenn der Midfoot nach dorsal (fußrückenwärts) beugt, der Hacken vom Erdboden abgehoben ist und der Vorfuß noch Kontakt mit dem Erdboden hat (Abb. 2).
Im Gegensatz zum Schimpansen besitzt der Mensch nach klassischer Vorstellung keinen mobilen Midfoot, sondern einen starren Midfoot mit Längsfußgewölbe. Dieses wird als wichtige Struktur für ein effizientes Gehen auf zwei Beinen angesehen. Das Auftreten des Längsfußgewölbes mit Verlust der midtarsalen Beweglichkeit wird deshalb als ein Schlüsselereignis im Rahmen der Entstehung des zweibeinigen menschlichen Ganges interpretiert.
Folgerichtig versuchen Paläanthropologen, knöcherne Merkmale bei frühen fossilen Homininen* zu identifizieren, die auf die Entstehung des Längsfußgewölbes und den Verlust der Midfoot-Beweglichkeit hinweisen. Es wurden jedoch schon einige Studien publiziert, die diese Vorstellungen stark in Frage stellen (Übersicht bei Brandt 2014). Greiner & Ball haben nun im American Journal of Physical Anthropology 2014 eine weitere interessante Untersuchung zu diesem Thema vorgelegt.
In der Studie von Greiner & Ball (2014) wurden an Fußknochen von menschlichen Leichen und Kadavern von Schimpansen, Pavianen und Makaken Anheftungen („pins“) vorgenommen und die Lage dieser Anheftungen während Beuge-Streck-Bewegungen überwacht. Das Schienbein wurde dabei von den Forschern über den Fuß bewegt. Es wurden die Flexion-Extensions-Muster und die damit verbundene Ausrichtung der Drehachsen des talonavicularen, calcaneocuboidalen und lateralen cubometatarsalen Gelenkes* (Abb. 1) untersucht.

Abb. 2: Rechter Fuß des Schimpansen in der Ansicht von außen mit midtarsalem „Bruch“ (Pfeil). (Nach Susman 1983)
Die Ergebnisse von Greiner & Ball (2014) zeigen, dass die Midfoot-Beweglichkeit primär im talonavicularen und lateralen cubometatarsalen Gelenk und nicht im calcaneocuboidalen Gelenk stattfindet. Noch überraschender ist jedoch, dass sowohl das Ausmaß der Bewegung als auch die Ausrichtung der Gelenkachsen bei Schimpanse und Mensch ähnlich sind. Welche funktionellen Konsequenzen ergeben sich daraus für Mensch und Schimpanse und welche Bedeutung haben diese Ergebnisse für die Interpretation fossiler homininer Fußknochen?
Entgegen klassischer Vorstellungen ist sowohl beim Menschen als auch beim Schimpansen das Potential für einen midtarsalen „Bruch“ und für einen „Sperrmechanismus“ zur Erhöhung der Fußsteife im Midfoot vorhanden. Die Fähigkeit zum midtarsalen „Bruch“ nicht nur beim Schimpansen, sondern auch beim Menschen ergibt sich aus der ähnlichen Beuge- und Streckfähigkeit im talonavicularen und lateralen cubometatarsalen Gelenk.
Beide Gelenke zeigen bei Mensch und Schimpanse auch eine ähnlich divergierende Ausrichtung, die einen „Sperrmechanismus“ im Midfoot bewirkt. Dieser „Sperrmechanismus“ verwandelt den menschlichen Fuß in einen steifen Hebel, der beim zweibeinigen Gehen ein effektives Abstoßen des Fußes vom Erdboden bewirkt. Dieser Mechanismus wird seit Jahrzehnten als wichtige Entwicklung im Rahmen der Entstehung des menschlichen zweibeinigen Ganges angesehen. Der „Sperrmechanismus“ wird mit Zunahme der Divergenz der Ausrichtung der Gelenkachsen effektiver, da mit größerer Divergenz die Wahrscheinlichkeit einer simultanen Beugung-Streckung des medialen und lateralen Fußes geringer wird. Die unterschiedliche Achsenausrichtung des Innen- und Außenfußes wird als verantwortlich für die Verhinderung einer größeren Beweglichkeit im talonavicularen und lateralen cubometatarsalen Gelenk während der Fußabdruckphase angesehen, auch wenn jedes Gelenk für sich allein stärker mobil ist.
Überraschend ist, dass Schimpansen nicht nur eine ähnlich divergierende Ausrichtung der inneren talonavicularen zur äußeren cubometatarsalen Gelenkachse zeigen, sondern dass die Variabilität der Ausrichtung sogar geringer als beim Menschen ist. Greiner & Ball (2014) wollen daraus allerdings nicht schließen, dass der „Sperrmechanismus“ des Midfoot beim Schimpansen noch effektiver als beim Menschen ist.
Greiner & Ball (2014) haben mit ihrer Untersuchung nachgewiesen, dass der Midfoot von Mensch und Schimpanse trotz unterschiedlicher Morphologie eine ähnliche Flexibilität mit der Fähigkeit zum midtarsalen „Bruch“ und zur Fußversteifung besitzt. Diese Fähigkeiten werden bei Mensch und Schimpanse im Leben aber in verschiedener Weise in Anspruch genommen, denn sie praktizieren eine sehr unterschiedliche Fortbewegung.
Die Studienergebnisse von Greiner & Ball (2014) haben große Konsequenzen für die Interpretation der Fußknochen fossiler Homininen. Entgegen z. B. Lovejoy et al. (2009), Ward et al. (2011), Zipfel et al. (2011) und Proctor (2013) können auf der Basis der Knochenmerkmale des Fußes früher fossiler Homininen keine Schlüsse auf die Fußbeweglichkeit und auf ein äußeres Längsfußgewölbe und damit auf bipede Fähigkeiten gezogen werden (siehe auch Brandt 2014).
Glossar
frühe Homininen: Übergangsformen zwischen großaffenähnlichen und menschlichen Wesen („Vormenschen“ oder „Affenmenschen“). Midfoot oder midtarsaler Fuß (vorderer Fußwurzelbereich): Os cuboideum, Os naviculare, Ossa cuneiformia. talonaviculares Gelenk: Gelenk zwischen dem Talus und dem Os naviculare. calcaneocuboidales Gelenk: Gelenk zwischen dem Calcaneus und dem Os cuboideum. laterales cubometatarsales Gelenk: Gelenk zwischen dem Os cuboideum und dem Os metatarsale 5.
M. Brandt
[Brandt M (2014) Wie sicher sind Deutungen in der Paläanthropologie? Australopithecus sediba und sein merkwürdiges Merkmalsmosaik. W+W Special Paper B-14-3 ; www.wort-und-wissen.de/artikel/sp/b-14-3-sediba.pdf (Zugriff am 20.4.2015) • Greiner TM & Ball KA (2014) Kinematics of primate midfoot flexibility. Am. J. Phys. Anthropol. 155, 610-620 • Lovejoy CO, Latimer B, Suwa G, Asfaw B & White TD (2009) Combining prehension and propulsion: the foot of Ardipithecus ramidus. Science 326, 72e1-72e8 • Proctor DJ (2013) Proximal metatarsal articular surface shape and the evolution of a rigid lateral foot in hominins. Journal of Human Evolution 65, 761-769 • Susman RL (1983) Evolution of the human foot: evidence from Plio-Pleistocene hominids. Foot & Ankle 3, 365-376 • Ward CV, Kimbel WH & Johanson DC (2011) Complete fourth metatarsal and arches in the foot of Australopithecus afarensis. Science 331, 750-753 • Zipfel B, DeSilva JM, Kidd RS, Carlson KJ, Churchill SE & Berger LR (2011) The foot and ankle of Australopithecus sediba. Science 333, 1417-1420.]