Moleküle und Anatomie im Konflikt? Die Systematik der Säugetiere
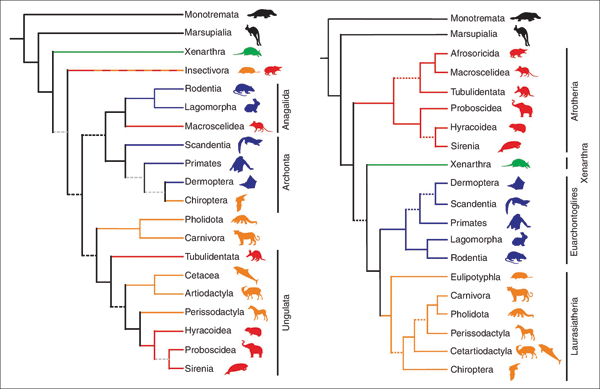
Abb. 1: Links der bislang akzeptierte Baum der plazentalen Säugetiere nach morphologischen Merkmalen, rechts der sich abzeichnende Baum nach molekularen Daten. (Aus Springer et al. 2004)
Die klassische evolutionsbiologische Sicht der Säugetierevolution lässt sich wie folgt zusammenfassen: Während der Zeit der Dinosaurier (Mesozoikum) existierten nur einzelne insektenjagende Vertreter urtümlicher Säugetiergruppen, die sich deutlich von den heutigen unterschieden. Der Einschlag eines großen Asteroiden vor ca. 66 Millionen Jahren schuf u. a. durch die darauf folgende weltweite Umweltzerstörung, durch den Klimawandel und das Aussterben der Dinosaurier den Freiraum für den Siegeszug der modernen plazentabesitzenden Säugetiere (Plazentalier). Innerhalb nur weniger Jahrmillionen, so die Geschichte weiter, traten zahlreiche Vertreter der modernen Ordnungen dieser Säugetiergruppe explosionsartig auf, begleitet von der ebenso rasch aufblühenden Vielfalt unter den Beuteltieren (Marsupialier), die einen anderen modernen Zweig der Säugetiere repräsentieren. Die klassische biologische Systematik der Säugetiere und die Fossilüberlieferung stützte dieses Bild bis zum Ende des 20. Jahrhunderts.
In den letzten zwanzig Jahren haben zahlreiche neue Fossilfunde und ein nahezu unüberschaubarer Zuwachs molekulargenetischer Daten das klassische Bild zur Evolution der Säugetiere jedoch aufgelöst. Tiefgreifende Änderungen bisheriger Modelle zur Systematik und Evolution der Säuger wurden vollzogen, andere sind nur teilweise akzeptiert oder werden unvermindert kontrovers diskutiert (Yoder et al. 2013).
Die bis zur Jahrtausendwende geltende Systematik der plazentalen Säugetiere unterschied auf der Grundlage morphologischer Kriterien die basale Überordnung die Xenarthra („Nebengelenktiere“ wie Ameisenbär, Faultier und Gürteltiere) von den Überordnungen der Ungulata (Huftiere, u. a. mit Seekühen, Rüsseltieren, Unpaarhufern, Paarhufern), der Archonta (u. a. mit Primaten, Fledermäusen, Spitzhörnchen) und der Anagalida (wie Nagetiere, Hasenartige, Spitzmaus) (Abb. 1, links). Die Integration von molekularen Daten änderte diese Gruppierung tiefgreifend. Der auf molekulargenetischen Daten basierende Stammbaum, den Springer et al. (2004) vorstellten, ordnet die 18 plazentalen Säugetierordnungen ebenfalls vier Überordnungen zu: Xenarthra, Afrotheria, Laurasiatheria und Euarchontoglires. Die zuletzt genannten drei neu geschaffenen Überordnungen tragen ihren Namen aufgrund der Region, in der ihre stammesgeschichtliche Abspaltung von den übrigen Überordnungen mit den ihnen zugeordneten Säugetierordnungen und -familien vermutet wird. Diese neuen Überordnungen fassen z. T. Ordnungen plazentaler Säugetiere auf eine Weise zusammen, wie sie nach morphologischen Kriterien nie zusammengestellt würden (Abb. 1, rechts). Z. B. stehen die Fledermäuse nun den Huftieren näher als den Primaten, und die Elefantenspitzmäuse (Rüsselspringer) gehören nicht mehr zu den Insektenfressern oder Hasenartigen, sondern sind eine Untergruppe der Afrotheria und damit enger verwandt mit den Erdferkeln.

Abb. 2: Madagassischer (links) und europäischer Igel. Eines von vielen Konvergenzbeispielen unter Säugetieren. (GNU Freie Dokumentations-Lizenz)
Nach Bininda Emonds et al. (2012) haben die vergleichend systematischen Forschungen der letzten Jahre dennoch eine weitestgehende Übereinstimmung von morphologischen und molekularbiologischen Datensätzen innerhalb der Xenarthra, Laurasiatheria und Euarchontoglires gezeigt (allerdings nur um den „Preis“ der Annahme vieler Konvergenzen, wenn diese Systematik stammesgeschichtlich interpretiert wird, s. u.). Bei den Afrotheria ist dies jedoch nicht gelungen. Der stammesgeschichtliche Ursprung dieser äußerlich sehr inhomogen erscheinenden Gruppe, zu der z. B. Seekühe, Erdferkel, Elefanten und die Goldmulle zählen, wird in Afrika vermutet.
Aufgrund der auch durch jüngere DNA-Sequenzanalysen gefestigten, modernen Systematik muss davon ausgegangen werden, dass im Rahmen der evolutionären Diversifikation der Säugetiere innerhalb und zwischen den einzelnen Überordnungen zahlreiche und ausgeprägte konvergente Merkmalsentwicklungen erfolgt sein müssen (z. B. Konvergenz zwischen madagassischem und europäischem Igel, Abb. 2). Die extrem hohe Anzahl von Konvergenzen, die bei den plazentalen Säugetieren, ob rezent oder fossil) anzunehmen ist, bezeichneten Springer et al. als Phänomen der korrelierenden Merkmalsevolution. Springer et al. (2004) forderten deshalb, dass künftige phylogenetische Modellierungen das Phänomen der korrelierenden Merkmalsevolution berücksichtigen müssen. Eine befriedigende evolutionstheoretische Erklärung dieses Phänomens, das laufend durch neue Befunde bestätigt wird, steht immer noch aus (vgl. dazu auch Ullrich 2012).
H. Ullrich
[Bininda-Emonds ORP et al. (2007) The delayed rise of present-day mammals. Nature 446, 507-512; Bininda-Emonds ORP et al. (2012) Rocking clocks and clocking rocks: a critical look at divergence time estimation in mammals. In: Asher JR & Müller J (eds) From Clone to Bone - The Synergy of Morphological and Molecular Tools in Palaeobiology. Cambridge, pp. 38-82; Yoder AD et al. (2013) Fossils versus Clocks. Science 339, 656-658; O’Leary MA et al. (2013) The Placental Mammal Ancestor and the post-K-Pg Radiation of Placentals. Science 339, 662-667; Springer MS et al. (2004) Molecules consolidate the placental mammal tree. Trends Ecol. Evol. 19, 430-438; Ullrich H (2012) Verwirrende Konvergenzen. Zur Evolution des Mittelohres der Säugetiere. Stud. Int. J. 19, 20-33.]