Pflanzliche Wiederbesiedlung in der Polarregion
Die Polarregionen bieten unwirtliche Lebensbedingungen und sind daher artenarm. Infolge der Klimaerwärmung und den daraus resultierenden besseren Lebensbedingungen wird die Einwanderung von neuen Pflanzen- und Tierarten erwartet. Welche Faktoren bestimmen die Einwanderung?
Die Inselgruppe um Spitzbergen (= Svalbard-Archipel) eignet sich gut für die Untersuchung der Geschwindigkeit von Migrationsvorgängen (Einwanderungen), da sie während der letzten Eiszeit vor 20 000 Jahren konventioneller Zeitrechnung vollständig von Eis bedeckt war, und sehr wahrscheinlich die gesamte Flora seitdem neu einwanderte. Svalbard liegt nördlich des Polarkreises und weit entfernt von potentiellen Ausgangsregionen der Wiederbesiedlung (Grönland, Nordrussland, Nordskandinavien). Seine Tundrenvegetation zählt nur etwa 130 verschiedene Blütenpflanzenarten. Eine Forschergruppe (Alsos et al. 2007) sammelte 4439 Exemplare von neun arktischen Blütenpflanzenarten sowohl auf Svalbard als auch in den genannten möglichen Quellregionen. Die neun Arten aus sechs Familien wurden so ausgewählt, dass sie hinsichtlich ihrer ökologischen Ansprüche für die Flora Svalbards repräsentativ sind. Drei Arten haben gefiederte Ausbreitungseinheiten (Samen bzw. Früchte) für die Windverbreitung, weitere drei fleischige Früchte für die Verbreitung durch Vögel und Säugetiere und die restlichen drei sind hinsichtlich der Verbreitung nicht spezialisiert.
Durch Vergleich des „genetischen Fingerabdruckes“ aller 4439 Exemplare wurde für jede Art ermittelt, aus welcher Ausgangsregion sie den Svalbard-Archipel kolonisierte. Außerdem wurde abgeschätzt, wieviele verschiedene Samen jeder Art mindestens nach Svalbard gelangt sein mussten, um die dort vorgefundene genetische Vielfalt zu erklären.
Die hauptsächliche Besiedlung durch die untersuchten neun Arten nach der Eiszeit erfolgte aus Nordrussland, mit Abstand gefolgt von Nordskandinavien und Grönland. Das dürfte damit zusammenhängen, dass zwischen Svalbard und Nordrussland im Gegensatz zu Skandinavien im Winter eine Verbindung durch verdriftende Eisschollen besteht, mit deren Hilfe die Ausbreitungseinheiten gut transportiert werden können. Bei mehreren Arten waren die in Svalbard nachgewiesenen Populationen unterschiedlich und konnten entsprechend auch unterschiedlichen Herkünften zugewiesen werden. Beispielsweise gehen die Vorkommen der Rauschbeere (Vaccinium uliginosum) sowohl auf russische, als auch grönländische Quellen zurück, während etwa die Krähenbeere (Empetrum nigrum) nur aus Grönland und die Moltebeere (Rubus chamaemorus) nur aus Nordrussland stammen. Für die Alpen-Gänsekresse (Arabis alpina) konnte keine Entscheidung getroffen werden, da diese Art in den fraglichen Gebieten genetisch einheitlich ist.
Bei den übrigen Arten wurde abgeschätzt, dass jeweils mehrere Besiedlungen stattgefunden haben mussten, indem im Falle der Moltebeere mindestens 6, bei der Silberwurz (Dryas octopetala) sogar 38 Ausbreitungseinheiten unabhängig voneinander zu den Inseln Svalbards gelangten und sich dort erfolgeich etablieren konnten. Wegen der anzunehmenden fehlgeschlagenen Versuche dürften in Wirklichkeit viel mehr Ausbreitungseinheiten nach Svalbard gelangt sein.
Bei 6 Arten legte der genetische Befund neben der hauptsächlichen Quellregion noch eine zweite Ausgangsregion nahe: Für die vier kälteresistentesten Arten D. octopetala, S. herbacea, C. tetragona und S. rivularis sowie für die beiden am meisten wärmeliebenden (und folglich seltensten) Arten B. und V. uliginosum.
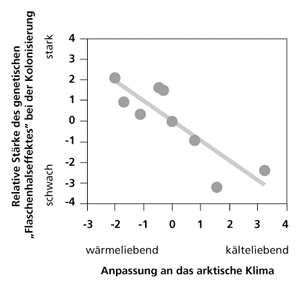
Abb.1: Zusammenhang zwischen der Anpassung an das Klima und der genetischen Vielfalt bei den Pflanzen auf Svalbard. Nach Alsos et al. (2007)
Die genetische Variabilität der untersuchten Pflanzen auf Svalbard korrelierte negativ mit ihrer Anpassung an das gegenwärtige dortige Klima: je besser eine Pflanzenart das Klima vertrug, desto größer war ihre genetische Vielfalt und umgekehrt (Abb. 1).
Nur von Juni bis August überschreiten die Mitteltemperaturen den Gefrierpunkt mit einem Maximum von 4,4 °C im Juli, und die Küstenregionen sind nur im Sommer für 6 Wochen eisfrei. Die Vegetation wird ermöglicht durch den Golfstrom, der während der 4-monatigen Polarnacht gegenüber dem auf dieser Breite üblichen Klima für relativ milde Winter sorgt.
Die Autoren kommen zu dem Schluss, dass bestimmend für die Neubesiedlung die ökologischen Bedingungen am Zielort sind und nicht die (offenbar immer in genügender Menge erfolgende) Verdriftung der Samen (durch Wind, Vögel oder z.B. Treibholz und -eis und selbst durch Säugetiere). Dieser Schluss wird unterstützt durch den Befund, dass 80 bis 90% der kälteresistenten Pflanzen in den potentiellen Ausgangsregionen auch auf Svalbard präsent sind, aber nur 40 bis 60% der relativ wärmeliebenden. Und damit würde ein wärmeres Klima auf Svalbard auch eine größere pflanzliche Vielfalt infolge erfolgreicher Migration erwarten lassen. Die gute Umweltresistenz bestimmter Pflanzensamen ist bekannt (Brown 2001, vgl. Lindemann 2001) und es können sich Pflanzen binnen weniger Jahre nicht nur vom Festland auf Inseln ausbreiten, sondern sich sogar mikroevolutiv an das neue Habitat anpassen (Cody et al. 1996, vgl. Kutzelnigg 1997). Auf der Basis der konventionellen Zeitrechnung veranschlagen Alsos et al. im Online support (www.sciencemag.org/cgi/content/full/316/5831/1606/DC1) 10 000 Jahre seit Beginn der Wiederbesiedlung. Aber auch wenn nur einige Prozent der verdrifteten Samen sich erfolgreich ansiedeln könnten, so wären selbst für Dryas octopetala nur einige 1000 Samen nötig. Die Leistungsfähigkeit der Transportprozesse dürfte die von Alsos et al. geschätzte Verdriftung von etwa einer Ausbreitungseinheit alle 2 Jahre bei weitem übersteigen (was einer erfolgreichen Ansiedlung einer Pflanze nur einmal pro Vierteljahrtausend entspräche). Damit stellt diese Studie indirekt die Datierung der letzten Eiszeit in Frage: Wäre Svalbard schon so lange teilweise eisfrei, müsste eine höhere genetische Vielfalt bestehen.
Nach einer größeren Katastrophe wäre jedenfalls zu erwarten, dass verwüstete, aber klimatisch günstige Regionen von vielleicht nur wenigen Ausgangspunkten aus auch über große Entfernungen rasch – innerhalb von Jahren – und durch nur wenige – einige Dutzend – Exemplare jeder Art wiederbesiedelt werden können, und genau das erwartet und sieht man ja auch heute bei der Artenmigration infolge der globalen Erwärmung.
WL
[Alsos IG, Eidesen PB, Ehrich D, Skrede I, Westergaard K, Jacobsen GH, Landvik JY, Taberlet P & Brochmann C (2007) Frequent long-distance plant colonization in the changing arctic, Science 316, 1606-1609; Brown K (2001) Patience yields secrets of seed longevity, Science 291, 1884-1885; Cody ML & Overton JM (1996) Short-term evolution of reduced dispersal in island plant populations. J. Ecology 84, 53-61; Kutzelnigg H (1997) Schnelle Evolution der Samenverbreitung bei Pflanzenpopulationen auf küstennahen Inseln. Stud. Int. J. 4, 39-40; Lindemann WB (2001) Langlebige Pflanzensamen. Stud. Int. J. 8, 101.]