Konvergenzen oder komplexe Ausgangsform? Brachiopoden (Armfüßer) und Saurier
Als konvergente Strukturen werden Organe bezeichnet, die eine gleiche oder sehr ähnliche Funktion haben, aber unabhängig voneinander bei Organismengruppen auftreten, die nicht als (nahe) verwandt gelten. Darüber hinaus sind konvergente Strukturen oft im Aufbau untereinander so ähnlich, daß sie nach den bekannten Homologiekriterien (ohne Vorgabe evolutionstheoretischer Hypothesen) nicht von Homologien unterschieden werden können. Da solche bauplangleichen Konvergenzen durch fortschreitende Forschung immer zahlreicher werden, führt das immer häufiger zu erheblichen Merkmalskonflikten bei Stammbaumdarstellungen (Junker 2003). Auch stellt sich dadurch – besonders bei Schlüsselmerkmalen! – immer dringlicher die Frage nach dem „Wie“ ihrer evolutiven Entstehung (vgl. Junker 2005b). Hier werden zwei weitere Beispiele von Konvergenzen genannt, die bei so unterschiedlichen Lebewesengruppen wie Brachiopoden und Sauriern auftreten.
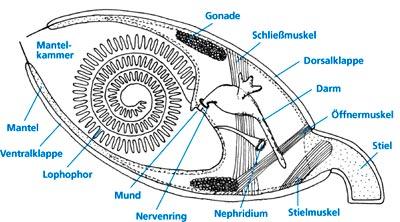
Abb.1: Senkrechter Schnitt durch einen Brachiopoden (Armfüßer; schematisch). Zwischen Dorsalklappe (Rückenklappe; oben) und Ventralklappe (Bauchklappe) der Weichkörper. Beherrschendes und namengebendes Organ sind zwei mit Tentakeln besetzte, hier spiralig aufgerollte Kiemenarme (Lobophor), davon ist einer dargestellt. (Nach Ziegler 1998).
Brachiopoden (Armfüßer) ähneln nur äußerlich Muscheln und sind jedem Fossiliensammler gut bekannt. Wie andere Tierstämme treten sie schon im Unterkambrium mit einer erstaunlichen Formenfülle auf (Jansen 2004); dieses für die meisten Tierstämme geltende Phänomen wird seit langem als „kambrische Explosion“ bezeichnet (Valentine 2004). Bis vor wenigen Jahrzehnten kannte man aus heutigen Meeren nur wenige noch existierende Brachipodenarten; sie gelten als „lebende Fossilien“ (Thenius 2000, 101-106). Die intensivierte Forschung hat jedoch inzwischen gezeigt, daß lebende Brachiopoden gar nicht so selten sind. Vielmehr sind ihre Diversität, Häufigkeit und Artenzahl viel größer als ursprünglich gedacht; inzwischen kennt man etwa 120 rezente Gattungen (Jansen 2004). Charakteristisch und namengebend für Armfüßer ist ein Paar fleischiger, mit Tentakeln besetzter Kiemenarme (Lobophor; Abb. 1). Damit erzeugt das Tier bei leicht geöffneten Schalenklappen einen Wasserstrom durch das Innere und fängt daraus mit den Kiemenarmen winzige Nahrungspartikel. Verschiedene Ordnungen fossiler Brachiopoden besaßen ein kalkiges, spiraliges Armgerüst (Atrypida, Athyridida und Spiriferida). Die Spiralkegel des Armgerüsts sind aber bei jeder Ordnung unterschiedlich ausgerichtet, wodurch der Filterstrom verschieden verläuft. Von diesen ausgestorbenen Gruppen wird angenommen, daß sie einst evolutiv aus der Gruppe der noch heute existierenden Rhynchonellida hervorgegangen sind. Für lebende Rhynchonelliden ist ein spiraliges Lobophor charakteristisch. Aufgrund seines wechselhaften fossilen Auftretens und seines verschiedenen Baues wird vermutet, „das spiralige Armgerüst könnte mehrfach unabhängig dadurch entstanden sein, daß entlang der spiralig aufgerollten Kiemenarme ein kalkiges Stützskelett abgeschieden wurde“ (Jansen 2004, 310).
Zahlreiche Funde von kleinen Dinosauriern mit federartigen Strukturen und echte Vögel aus der Unterkreide Chinas machen seit Anfang der 1990er Jahre Furore (Überblick bei Junker 2005a). Diese teils anscheinend wirklich federartigen, teils deutlich von Federn abweichenden Hautstrukturen treten aber nicht nur bei zweibeinigen Raubdinosauriern auf (Theropoden); diese gehören zur Großgruppe der Echsenbecken-Dinosaurier (Saurischia) und gelten aufgrund ihrer Skelettmerkmale nach der derzeitigen Mehrheitsmeinung als evolutiver Ursprung der Vögel (vgl. jedoch Junker 2005a, 55f.). Solche komplexen Hautanhänge werden aber auch von Papageienschnabel-Dinosauriern (Psittacosaurier) beschrieben (vgl. Mayr 2004, 27), die jedoch zur anderen Großgruppe, den Vogelbecken-Dinosauriern (Ornithischia), gehören. Letztere gelten heute – trotz des vogelähnlichen Beckens – aufgrund der Gesamtheit ihrer Skelettmerkmale nicht als Ursprungsgruppe der Vögel. Im Zusammenhang mit dem Neufund eines ca. 1 m langen Psittacosauriers aus China, der etwa 100 rätselhafte, borstenartige Strukturen von jeweils 24 cm Länge auf der Oberseite seines Schwanzes trägt, meint Mayr (2004, 27f), daß eine „haarartige Körperbedeckung“ durchaus mehrfach unabhängig voneinander entstanden sein“ könnte (vgl. den Beitrag von M. Stephan über Flugsaurier S. 86 ff. in dieser Ausgabe). Denkbar sei „aber auch, daß sie schon bei den gemeinsamen Vorfahren von Flugsauriern, Psittacosauriern und Vögeln vorhanden war, jedoch bei Krokodilen, Squamaten (zu denen die Brückenechse sowie Eidechsen, Warane und Schlangen gehören) und vielleicht auch Schildkröten (deren genaue Verwandtschaftsbeziehungen noch unsicher sind) sekundär wieder verloren ging“.
Doch geben die (bisherigen) Daten keine Basis für solche weitreichenden Spekulationen; sie würde darauf hinauslaufen, die komplexere Konstellation als ursprünglich zu betrachten. Wenn zumindest ein Teil der (komplexen) Hautstrukturen bei verschiedenen reptilhaften Wirbeltiergruppen sich abschließend als Federn bestätigen (sollten), bedeutet(e) dieser konvergente Befund eine Schwächung der Abstammungshypothese „zweibeinige Raubdinosaurier-Vögel“, weil dieses Merkmal an „Beweiskraft“ verlieren würde.
MS
[Jansen U (2004) Armfüßer – von der „kambrischen Explosion“ bis heute. Nat. u. Mus. 134, 304-314; Junker R (2003) . Stud. Int. J. 10, 3-11; Junker R (2005a) . Stud. Int. J. 12, 51-57; Junker R (2005b) . Stud. Int. J. 12, 74-76; Mayr G (2004) Ein Papageienschnabel-Dinosaurier mit „Borsten“ am Schwanz. Nat. u. Mus. 134, 26-28; Thenius E (2000) Lebende Fossilien. München; Valentine JW (2004) On the Origin of Phyla. Chicago and London; Ziegler B (1998) Spezielle Paläontologie. Stuttgart.]