
Modernes Mittelohr bei alten Säugetieren
Downloads
Das Mittelohr mit den Gehörknöchelchen Hammer, Amboss und Steigbügel repräsentiert eines der markantesten Merkmale der Säugetiere. Ihre Evolution aus Kiefergelenkknochen säugetierähnlicher Reptilien galt lange Zeit als Paradebeispiel für eine lineare und einmalige evolutionäre Umwandlung. In den letzten 25 Jahren zeigten zahlreiche fossile Funde einen unerwarteten Merkmalsmix innerhalb der frühen Säugetierformen, was zu einem weitreichenden Umdenken bezüglich des hypothetischen Evolutionsverlaufs bei Säugetieren führte. Diese Situation wird durch neuere Funde wie Microtherulum noch verwirrender.
Säugetiere repräsentieren eine vielfältige Tierklasse. Man kennt heute ca. 6.600 lebende Säugetierarten sowie fossile Vertreter von mehr als 4.000 Gattungen. Neben den aktuell lebenden Säugergruppen (Plazentatiere, Beuteltiere und Kloakentiere) finden sich ca. 20 Säugetierlinien in der fossilen Überlieferung. Die heutigen Hauptgruppen der Säuger (Kronen-Säugetiere) sind bereits im Mesozoikum (Erdmittelalter ca. 251 bis 66 Millionen radiometrische Jahre [MrJ]) ausdifferenziert. Den aus diesem geologischen Abschnitt bekannten ca. 540 Dinosauriergattungen stehen mittlerweile ca. 310 Säugetiergattungen gegenüber, zwei Drittel davon wurden erst in den letzten 25 Jahren entdeckt (Luo 2007 a; 2007 b).
Klassische Modelle der Mittelohrevolution
Ein besonderes Kennzeichen (Schlüsselmerkmal) der Säugetiere stellt das Mittelohr mit den charakteristischen drei Mittelohrknochen (MOK; vgl. Abb. 1) dar: Hammer (Malleus), Amboss (Incus) und Steigbügel (Stapes). Deren Evolution galt bis zum Ende des 20. Jahrhunderts als Paradebeispiel für eine lineare evolutionäre Umwandlung (z. B. in Form der Reichert-Gauppschen-Theorie, vgl. Ullrich 1994). Bis vor 25 Jahren ging man davon aus, dass die Evolution der Säugetiere einmalig (monophyletisch), stufenweise und kontinuierlich (graduell) ohne Sprünge oder rasante Diversifikationen erfolgte. Das bedeutet: Auch die Evolution der Umwandlung von Kieferknochen der Reptilien (z. B. Quadratum, Artikulare, Angulare) in die MOK der Säugetiere verlief graduell und monophyletisch (vgl. Abb. 5).
Figure 1. Abb. 1 Mittelohrknöchelchen des Menschen. (Nach Bertolini & Leutert 1982)
Die ersten Schritte sah man in frühen Therapsiden (säugetierähnlichen Reptilien) verwirklicht, die eine etwas kleinere Anlage der hinteren (postdentalen) Kieferknochen zeigten. Der zahntragende Knochen (Dentale) dehnte sich – so die Theorie – nach hinten und hinten oben aus und etablierte so ein sekundäres Kiefergelenk mit der Schläfenbeinschuppe des Schädels (Squamosum): Das Squamoso-Dental-Gelenk. Ein weiterer hinterer Kiefergelenkknochen, das Angulare, entwickelte eine nach hinten orientierte knöcherne Ausziehung (z. B. beim Dimetrodon-Pelycosaurier = „reflected lamina“), welcher eventuell die Basis des ringförmigen Knochens der frühen Säuger dargestellt haben könnte, der das Trommelfell aufspannt (Ectotympanon).
Figure 2. Abb. 2 Systematik der Säugetiere im weiteren Sinne (Mammaliaformes). Dunkelblau: Schwestergruppen der Säugetiere im engeren Sinne (Mammalia). Gelb: ausgestorbene Gruppen von Kronen-Säugetieren. Grün: Kloakentiere (Monotremata). Hellblau: Beutelsäuger (Metatheria/Marsupialia). Rot: höhere Säugetiere oder Plazentatiere (Eutheria/Plazentalia). (Nach Luo 2007)
Einige höhere Cynodonta (artenreichste Gruppe innerhalb der Therapsiden) zeigen fossil ein doppelt gelagertes Kiefergelenk (wie Diarthrognathus). Diese Situation wurde als ein Zwischenstadium gedeutet: Das seitlich gelegene Squamoso-Dental-Gelenk übernimmt mehr und mehr die Kaufunktion und entlässt die innen gelegenen primären postdentalen Kieferelemente (Quadratum, Artikulare, Angulare) aus dem Kiefergelenk, sodass sie später in das Mittelohr wandern. Die anatomische Situation bei Morganuconodon wurde als Prototyp für den Start der Entwicklung des MME („mammalian middle ear“) zum DMME „definitive mammalian middle ear“ betrachtet, da hier die postdentalen Kieferelemente (Angulare, Artikulare, Präartikulare und Quadratum) ausschließlich Strukturen des Hörapparates sind – vollständig getrennt vom Kauapparat (Meng et al. 2011). Spätere Autoren führten unter Berücksichtigung zahlreicher Fossilfunde eine neue Nomenklatur für die Mittelohrentwicklung bei Säugetieren ein. Mittlerweile unterscheidet man in der Evolutionsbiologie drei evolutionäre bzw. phylogenetische Stufen in der Entwicklung des Säugetiermittelohrs: das postdentale Mittelohr (postdentary-attached middle ear: PAME), das meckelianische Mittelohr (Meckelian-attached middle ear: MAME) und das vollständig vom Unterkiefer freie Mittelohr (detached middle ear: DME; s. Luo 2011; Meng et al. 2011; Wang et al. 2021; vgl. dazu Abb. 3).
Figure 3. Abb. 3 Holotypus (IVPP 24190) des unterkreidezeitlichen Säugetiers (Eutheria) Microtherulum oneirodes. (Wang & Wang 2023, CC BY 4.0 Deed, https://creativecommons.org/licenses/by/4.0/)
Phänomen der korrelierenden Merkmalsevolution
War man gegen Ausgang des 20. Jahrhunderts noch überzeugt, die bis dahin geltenden evolutionären Rekonstruktionen durch Fossilien bestätigt zu sehen (Luo et al. 2001), änderte sich dieses optimistische Bild dramatisch in den letzten 25 Jahren. Zahlreiche neue fossile Funde zeigten einen unerwarteten Merkmalsmix, der gemeinsam mit Daten aus der Embryologie und der Molekularbiologie zu einem weitreichenden Umdenken bezüglich des hypothetischen Evolutionsverlaufs bei Säugetieren zwang (Ullrich 2009, 2012, 2015): Das Modell einer monophyletischen, graduellen Evolution ohne Sprünge musste aufgegeben werden. Aufgrund der auch durch jüngere DNA-Sequenzanalysen etablierten Systematik bleibt nur der Schluss, dass im Rahmen der evolutionären Aufspaltung (Diversifikation) der Säugetiere innerhalb und zwischen den einzelnen Großgruppen zahlreiche und hochgradig konvergente (mehrfach unabhängig erworbene) Merkmalsentwicklungen erfolgt sein müssen (vgl. Abb. 5 sowie Ullrich 2013). Die extrem hohe Anzahl anzunehmender Konvergenzen, die bei den heutigen und fossilen plazentalen Säugetieren unter Vorgabe einer abgelaufenen Evolution anzunehmen ist, bezeichneten Springer et al. (2004) als Phänomen der korrelierenden Merkmalsevolution.
Zahlreiche neue fossile Funde zeigten einen unerwarteten Merkmalsmix und erzwangen ein Umdenken.
Auch Luo (2011) stellt fest: „Die Evolution der frühesten Säugetiere zeigt aufeinander folgende Episoden von Diversifikationen. Die Aufteilung in unterschiedliche Linien bei mesozoischen Säugetieren ist gekoppelt an viele unabhängige evolutionäre Experimente und ökologische Spezialisierungen.“ Diese Einschätzung gilt damit auch für die Evolution des Mittelohres bei unterschiedlichen Säugetiergruppen. Das Phänomen der korrelierenden Merkmalsevolution bleibt auch weiterhin trotz oder gerade wegen der Verfügbarkeit einer sehr großen Anzahl fossiler Dokumente von Säugetieren aus Erdmittelalter und -neuzeit hinsichtlich der verursachenden Mechanismen rätselhaft und unverstanden. Trägt vielleicht ein neuer fossiler Fund zur Klärung der Situation bei?
Microtherulum oneirodes – Ein traumhaftes Fossil
2023 stellten Wang & Wang die fossilen Überreste eines fast vollständig erhaltenen, ca. 10 cm großen, modernen, höheren Säugetiers (Eutheria) aus der Unterkreide (118–120 radiometrische Mio. Jahre) unter dem Namen Microtherulum oneirodesvor (Abb. 3). Der Name „Micro“ bezieht sich auf die sogenannte mikrotypische Konfiguration des Mittelohrs, die charakteristisch für heutige Mäuse und Fledermäuse ist und auf ein hochfrequentes Hören schließen lässt (vgl. Abb. 4: Bauplantypen des Hammer-Amboss-Gelenkes moderner Säugetiere). „Therulum“ bedeutet„kleines Tierchen“ und „oneirodes“ (griechisch) steht für „traumhaft“. Die Autoren sehen in Microtherulum oneirodes einen traumhaften Fossilfund, welcher die Lücke in der frühen Mittelohr-Evolution der modernen Beutel- und Plazenta-Säugetiere (Theria) endlich füllen soll. Insbesondere waren die Erwartungen auch deshalb so groß, weil die fossilisierten knöchernen Elemente des Mittelohres bei Microtherulum oneirodes hervorragend erhalten waren. Microtherulum oneirodeszeigt eine sattelförmige Gelenkverbindung zwischen dem Hammer und dem Amboss (sog. Incudomallear-Gelenk). Diese ist für eine große Gruppe moderner (rezenter) Säugetiere typisch (z. B. für Fleischfresser, Nagetiere, Fledermäuse; vgl. Abb. 4 über die Bauplantypen des Incudomallear-Gelenkes bei lebenden Säugetieren).
Microtherulum stellt den frühesten bekannten fossilen Beleg für die Existenz eins modernen sattelartigen Hammer-Amboss-Gelenks dar.
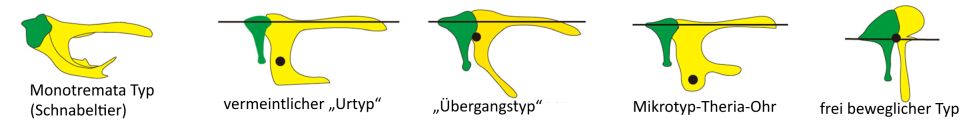
Figure 4. Abb. 4 Bauplantypen des Hammer-Amboss-Gelenkes in einer Ansicht von hinten bei (v. l. n. r.): Kloakentieren (Schnabeltier), Beuteltieren und Plazentatieren: vermeintlicher „Urtyp“ (Opossums), „Übergangstyp“ (Hamster), Mikrotyp-Theria-Ohr (Mäuse, Fledermäuse, Microtherulum oneirodes) und frei beweglicher Typ (Eichhörnchen). Bei jedem Bauplantyp zeigt der Punkt das geschätzte Massenzentrum der Gehörknöchelchenkette. (Nach Wang & Wang 2023, CC BY 4.0 Deed, http://creativecommons.org/licenses/by/4.0/)
Die Gelenkverbindung zwischen Hammer und Amboss von Microtherulum stellt den frühesten bekannten fossilen Beleg für die Existenz eines sattelartigen Hammer-Amboss-Gelenks dar. Daraus muss geschlussfolgert werden, dass diese für die modernen Säugetiere charakteristische Gelenkkonfiguration bereits bei ihren ältesten bekannten Vertretern (Eutheriern) in der unteren Kreide ausgebildet war.
Konvergenz und paralleles Auftreten der Mittelohrstrukturen
Wichtig zu erwähnen ist, dass diese moderne Konfiguration des Mittelohres bei Microtherulum oneirodeszeitgleich zu anderen Formvarianten bei den Schwestergruppen der modernen Säugetiere existierte (Haramiyida, Multituberculata, Eutriconodonta, Zhangheotheriidae und Monotremata [Kloakentiere]). Damit ist es in biostratigrafischer Hinsicht nicht mehr möglich, von „primitiveren“ und „modernen“ Varianten der MOK zu sprechen. Ein häufig als primitiv gewertetes Merkmal bei diesen zeitgleich lebenden Schwestergruppen ist ein teilweise verknöcherter Meckelscher Knorpel wie auch eine spezielle Furche im Unterkiefer für den Meckelknorpel – und wird deshalb als morphologisches Bindeglied bei der Mittelohrentwicklung bei Säugetieren dargestellt. Diese Struktur fehlt beim zeitgleich lebenden Microtherulum oneirodesvollständig.Die Autoren Wang & Wang (2023) ziehen daraus den Schluss: „Es scheint, dass der Abbau des Meckelschen Knorpels auch bei den Eutheriern der frühen Kreidezeit variabel ist, was auf eine hohe evolutionäre Variabilität bei der Ablösung des Mittelohrs bei den frühen Säugetierformen mit Ausnahme der Allotherier hinweist.“
Aus evolutionärer Perspektive müssen vielfache, unabhängige und gleichgerichtete Entwicklungssprünge angenommen werden.
Abb. 5 dokumentiert die große Variabilität der Mittelohrformen bei den Säugetieren im weiteren Sinne (Mammaliaformes). Darüber hinaus wird erneut belegt, dass aus evolutionärer Perspektive vielfache, unabhängige und gleichgerichtete Entwicklungssprünge bei der Entstehung des Mittelohres angenommen werden müssen. Das freistehende Mittelohr (detached middle ear, DME) und die damit verbundene Entkopplung von Hör- und Kauapparat entwickelte sich nach der aktuellen Phylogenie mehrfach in mesozoischen Säugetierformen (Abb. 5). Als zugrundeliegender Mechanismus für das Entkopplungsereignis (wobei man hier richtiger von einem hypothetischen Selektionsvorteil sprechen sollte), wird von den Autoren lediglich die Anpassung an ein effizientes Kauen ins Spiel gebracht.
Figure 5. Abb. 5 Hypothetische Evolutionsstufen der Mittelohrregion in einer vereinfachten Phylogenie der Säugetiere inkl. der evolutionären Entwicklung zu den modernen tribosphenischen Backenzähnen (s. unten links: einfache Form: prätribosphenische Gruppe, Übergangsform: vielhöckrige Zahn-Gruppe, säugetiertypische Form: tribosphenische Gruppe), sowie der Abtrennung des Mittelohrs vom Unterkiefer durch die Umwandlung von Unterkieferknochen bei Reptilien (Quatratum, Artikulare, Angulare) zum Hammer-Amboss-Gelenk. Die verschiedenen Elemente des Mittelohrs sind markiert, das Ektotympanon in Lila, der Hammer (Malleus) in Gelb, der Amboss (Incus) in Grün und der verknöcherte Meckel‘sche Knorpel in Orange. Man beachte das konvergente, gleichzeitige und zum Teil rückschrittliche Auftreten der tribosphenischen bzw. der prätribosphenischen Formen, des DME, des MAME, eines verknöcherten Meckel‘schen Knorpels und des überlappenden Gelenkes zwischen Hammer und Amboss in den unterschiedlichen rezenten und ausgestorbenen Säugetiergruppen. (Nach Wang & Wang 2023, CC BY 4.0 Deed, http://creativecommons.org/licenses/by/4.0/).
Diskussion
Der aktuelle Fossilfund des modernen Säugetiers Microtherulum oneirodes aus der Unterkreide bestätigt, dass aus evolutionärer Perspektive davon ausgegangen werden muss, dass auch die Entstehung des Mittelohres und seiner Begleitstrukturen bei modernen Säugetieren und ihren fossilen Verwandten mehrfach und unabhängig voneinander erfolgt sein muss. Außerdem hilft das Fossil nicht aufzuklären, von welchen Vorfahren die für moderne Säugetiere typische Konfiguration der Gelenkverbindungen zwischen Hammer und Amboss (Incudomalleolar-Gelenk) abzuleiten ist, da diese in den frühesten Formen bereits völlig modern erscheint. Die zugrundeliegenden Mechanismen für die postulierten konvergenten Entwicklungen und die dafür erforderlichen Selektionsbedingungen der Mittelohrknochen bei Säugetieren bleiben auch nach dem Fund von Microtherulum oneirodes unbekannt. Der Fund von Microtherulum oneirodes ist ein erneutes Beispiel für das allgegenwärtige und kausal ungeklärte Phänomen der „korrelierenden“/konvergenten Merkmalsevolution in der Säugetierevolution. Die damit verbundene Untauglichkeit von Homologien (Merkmalsähnlichkeiten) bei der Rekonstruktion makroevolutionärer Vorgänge wird erneut exemplarisch bestätigt. Dieser Zusammenhang wiegt umso schwerer, da Homologien nach wie vor als das wesentlichste Argument für die gemeinsame Abstammung der Lebewesen aus dem Blickwinkel der Darwin‘schen Evolution gelten.
Literatur
Bertolini R & Leutert G (1982) Atlas der Anatomie des Menschen Nach systematischen und topographischen Gesichtspunkten. Band 3: Kopf und Hals, Gehirn, Rückenmark und Sinnesorgane. Leipzig.
Luo ZX et al. (2007 a) A new eutriconodont mammal and evolutionary development in early mammals. Nature 446, 288–293.
Luo, ZX. (2007 b) Transformation and diversification in early mammal evolution. Nature 450, 1011–1019.
Luo ZX (2011) Developmental patterns in Mesozoic evolution of mammal ears. Annu. Rev. Ecol., Evol. Syst. 42, 355–380.
Luo ZX, Crompton AW & Sun AL (2001) A new mammaliaform from the early Jurassic and evolution of mammalian characteristics. Science 292, 1535–1540.
Meng J, Wang Y & Li C (2011) Transitional mammalian middle ear from a new Cretaceous Jehol eutriconodont. Nature 472, 181–185.
Springer MS et al. (2004) Molecules consolidate the placental mammal tree. Trends Ecol. Evol. 19, 430–438.
Takechi & Kuratani (2010) History of studies on mammalian middle ear evolution. J. Exp. Zool. 314B, 417–433.
Ullrich H (1994) Embryologie und Homologie – Die Reichert-Gauppsche Theorie. Stud. Int. J. 1, 15–24.
Ullrich H (2009) Das Ende der klassischen Homologisierung. Neue Methoden in der vergleichenden embryonalen Forschung. Stud. Integr. J. 12, 58–65.
Ullrich H (2012) Verwirrende Konvergenzen. Zur Evolution des Mittelohres der Säugetiere. Stud. Integr. J. 19, 20–33.
Ullrich H (2013) Moleküle und Anatomie im Konflikt? Die Systematik der Säugetiere. Stud. Integr. J. 20, 119–121.
Ullrich H (2015) Wann entstanden die modernen Säugetiere? Einsichten aus Fossilien, Molekülen und Datierungen. Stud. Integr. J. 22, 23–30.
Wang H & Wang Y (2023) Middle ear innovation in Early Cretaceous eutherian mammals. Nat. Commun. 14:6831, 1–12.
Wang J. et al. (2021) A monotreme-like auditory apparatus in a Middle Jurassic haramiyidan. Nature 590, 279–283.
Das Mittelohr mit den Gehörknöchelchen Hammer, Amboss und Steigbügel repräsentiert eines der markantesten Merkmale der Säugetiere. Ihre Evolution aus Kiefergelenkknochen säugetierähnlicher Reptilien galt lange Zeit als Paradebeispiel für eine lineare und einmalige evolutionäre Umwandlung. In den letzten 25 Jahren zeigten zahlreiche fossile Funde einen unerwarteten Merkmalsmix innerhalb der frühen Säugetierformen, was zu einem weitreichenden Umdenken bezüglich des hypothetischen Evolutionsverlaufs bei Säugetieren führte. Diese Situation wird durch neuere Funde wie Microtherulum noch verwirrender.
Das Mittelohr mit den Gehörknöchelchen Hammer, Amboss und Steigbügel repräsentiert eines der markantesten Merkmale der Säugetiere. Ihre Evolution aus Kiefergelenkknochen säugetierähnlicher Reptilien galt lange Zeit als Paradebeispiel für eine lineare und einmalige evolutionäre Umwandlung. In den letzten 25 Jahren zeigten zahlreiche fossile Funde einen unerwarteten Merkmalsmix innerhalb der frühen Säugetierformen, was zu einem weitreichenden Umdenken bezüglich des hypothetischen Evolutionsverlaufs bei Säugetieren führte. Diese Situation wird durch neuere Funde wie Microtherulum noch verwirrender.
Säugetiere repräsentieren eine vielfältige Tierklasse. Man kennt heute ca. 6.600 lebende Säugetierarten sowie fossile Vertreter von mehr als 4.000 Gattungen. Neben den aktuell lebenden Säugergruppen (Plazentatiere, Beuteltiere und Kloakentiere) finden sich ca. 20 Säugetierlinien in der fossilen Überlieferung. Die heutigen Hauptgruppen der Säuger (Kronen-Säugetiere) sind bereits im Mesozoikum (Erdmittelalter ca. 251 bis 66 Millionen radiometrische Jahre [MrJ]) ausdifferenziert. Den aus diesem geologischen Abschnitt bekannten ca. 540 Dinosauriergattungen stehen mittlerweile ca. 310 Säugetiergattungen gegenüber, zwei Drittel davon wurden erst in den letzten 25 Jahren entdeckt (Luo 2007 a; 2007 b).
Klassische Modelle der Mittelohrevolution
Ein besonderes Kennzeichen (Schlüsselmerkmal) der Säugetiere stellt das Mittelohr mit den charakteristischen drei Mittelohrknochen (MOK; vgl. Abb. 1) dar: Hammer (Malleus), Amboss (Incus) und Steigbügel (Stapes). Deren Evolution galt bis zum Ende des 20. Jahrhunderts als Paradebeispiel für eine lineare evolutionäre Umwandlung (z. B. in Form der Reichert-Gauppschen-Theorie, vgl. Ullrich 1994). Bis vor 25 Jahren ging man davon aus, dass die Evolution der Säugetiere einmalig (monophyletisch), stufenweise und kontinuierlich (graduell) ohne Sprünge oder rasante Diversifikationen erfolgte. Das bedeutet: Auch die Evolution der Umwandlung von Kieferknochen der Reptilien (z. B. Quadratum, Artikulare, Angulare) in die MOK der Säugetiere verlief graduell und monophyletisch (vgl. Abb. 5).
Figure 1. Abb. 1 Mittelohrknöchelchen des Menschen. (Nach Bertolini & Leutert 1982)
Die ersten Schritte sah man in frühen Therapsiden (säugetierähnlichen Reptilien) verwirklicht, die eine etwas kleinere Anlage der hinteren (postdentalen) Kieferknochen zeigten. Der zahntragende Knochen (Dentale) dehnte sich – so die Theorie – nach hinten und hinten oben aus und etablierte so ein sekundäres Kiefergelenk mit der Schläfenbeinschuppe des Schädels (Squamosum): Das Squamoso-Dental-Gelenk. Ein weiterer hinterer Kiefergelenkknochen, das Angulare, entwickelte eine nach hinten orientierte knöcherne Ausziehung (z. B. beim Dimetrodon-Pelycosaurier = „reflected lamina“), welcher eventuell die Basis des ringförmigen Knochens der frühen Säuger dargestellt haben könnte, der das Trommelfell aufspannt (Ectotympanon).
Figure 2. Abb. 2 Systematik der Säugetiere im weiteren Sinne (Mammaliaformes). Dunkelblau: Schwestergruppen der Säugetiere im engeren Sinne (Mammalia). Gelb: ausgestorbene Gruppen von Kronen-Säugetieren. Grün: Kloakentiere (Monotremata). Hellblau: Beutelsäuger (Metatheria/Marsupialia). Rot: höhere Säugetiere oder Plazentatiere (Eutheria/Plazentalia). (Nach Luo 2007)
Einige höhere Cynodonta (artenreichste Gruppe innerhalb der Therapsiden) zeigen fossil ein doppelt gelagertes Kiefergelenk (wie Diarthrognathus). Diese Situation wurde als ein Zwischenstadium gedeutet: Das seitlich gelegene Squamoso-Dental-Gelenk übernimmt mehr und mehr die Kaufunktion und entlässt die innen gelegenen primären postdentalen Kieferelemente (Quadratum, Artikulare, Angulare) aus dem Kiefergelenk, sodass sie später in das Mittelohr wandern. Die anatomische Situation bei Morganuconodon wurde als Prototyp für den Start der Entwicklung des MME („mammalian middle ear“) zum DMME „definitive mammalian middle ear“ betrachtet, da hier die postdentalen Kieferelemente (Angulare, Artikulare, Präartikulare und Quadratum) ausschließlich Strukturen des Hörapparates sind – vollständig getrennt vom Kauapparat (Meng et al. 2011). Spätere Autoren führten unter Berücksichtigung zahlreicher Fossilfunde eine neue Nomenklatur für die Mittelohrentwicklung bei Säugetieren ein. Mittlerweile unterscheidet man in der Evolutionsbiologie drei evolutionäre bzw. phylogenetische Stufen in der Entwicklung des Säugetiermittelohrs: das postdentale Mittelohr (postdentary-attached middle ear: PAME), das meckelianische Mittelohr (Meckelian-attached middle ear: MAME) und das vollständig vom Unterkiefer freie Mittelohr (detached middle ear: DME; s. Luo 2011; Meng et al. 2011; Wang et al. 2021; vgl. dazu Abb. 3).
Figure 3. Abb. 3 Holotypus (IVPP 24190) des unterkreidezeitlichen Säugetiers (Eutheria) Microtherulum oneirodes. (Wang & Wang 2023, CC BY 4.0 Deed, https://creativecommons.org/licenses/by/4.0/)
Phänomen der korrelierenden Merkmalsevolution
War man gegen Ausgang des 20. Jahrhunderts noch überzeugt, die bis dahin geltenden evolutionären Rekonstruktionen durch Fossilien bestätigt zu sehen (Luo et al. 2001), änderte sich dieses optimistische Bild dramatisch in den letzten 25 Jahren. Zahlreiche neue fossile Funde zeigten einen unerwarteten Merkmalsmix, der gemeinsam mit Daten aus der Embryologie und der Molekularbiologie zu einem weitreichenden Umdenken bezüglich des hypothetischen Evolutionsverlaufs bei Säugetieren zwang (Ullrich 2009, 2012, 2015): Das Modell einer monophyletischen, graduellen Evolution ohne Sprünge musste aufgegeben werden. Aufgrund der auch durch jüngere DNA-Sequenzanalysen etablierten Systematik bleibt nur der Schluss, dass im Rahmen der evolutionären Aufspaltung (Diversifikation) der Säugetiere innerhalb und zwischen den einzelnen Großgruppen zahlreiche und hochgradig konvergente (mehrfach unabhängig erworbene) Merkmalsentwicklungen erfolgt sein müssen (vgl. Abb. 5 sowie Ullrich 2013). Die extrem hohe Anzahl anzunehmender Konvergenzen, die bei den heutigen und fossilen plazentalen Säugetieren unter Vorgabe einer abgelaufenen Evolution anzunehmen ist, bezeichneten Springer et al. (2004) als Phänomen der korrelierenden Merkmalsevolution.
Zahlreiche neue fossile Funde zeigten einen unerwarteten Merkmalsmix und erzwangen ein Umdenken.
Auch Luo (2011) stellt fest: „Die Evolution der frühesten Säugetiere zeigt aufeinander folgende Episoden von Diversifikationen. Die Aufteilung in unterschiedliche Linien bei mesozoischen Säugetieren ist gekoppelt an viele unabhängige evolutionäre Experimente und ökologische Spezialisierungen.“ Diese Einschätzung gilt damit auch für die Evolution des Mittelohres bei unterschiedlichen Säugetiergruppen. Das Phänomen der korrelierenden Merkmalsevolution bleibt auch weiterhin trotz oder gerade wegen der Verfügbarkeit einer sehr großen Anzahl fossiler Dokumente von Säugetieren aus Erdmittelalter und -neuzeit hinsichtlich der verursachenden Mechanismen rätselhaft und unverstanden. Trägt vielleicht ein neuer fossiler Fund zur Klärung der Situation bei?
Microtherulum oneirodes – Ein traumhaftes Fossil
2023 stellten Wang & Wang die fossilen Überreste eines fast vollständig erhaltenen, ca. 10 cm großen, modernen, höheren Säugetiers (Eutheria) aus der Unterkreide (118–120 radiometrische Mio. Jahre) unter dem Namen Microtherulum oneirodesvor (Abb. 3). Der Name „Micro“ bezieht sich auf die sogenannte mikrotypische Konfiguration des Mittelohrs, die charakteristisch für heutige Mäuse und Fledermäuse ist und auf ein hochfrequentes Hören schließen lässt (vgl. Abb. 4: Bauplantypen des Hammer-Amboss-Gelenkes moderner Säugetiere). „Therulum“ bedeutet„kleines Tierchen“ und „oneirodes“ (griechisch) steht für „traumhaft“. Die Autoren sehen in Microtherulum oneirodes einen traumhaften Fossilfund, welcher die Lücke in der frühen Mittelohr-Evolution der modernen Beutel- und Plazenta-Säugetiere (Theria) endlich füllen soll. Insbesondere waren die Erwartungen auch deshalb so groß, weil die fossilisierten knöchernen Elemente des Mittelohres bei Microtherulum oneirodes hervorragend erhalten waren. Microtherulum oneirodeszeigt eine sattelförmige Gelenkverbindung zwischen dem Hammer und dem Amboss (sog. Incudomallear-Gelenk). Diese ist für eine große Gruppe moderner (rezenter) Säugetiere typisch (z. B. für Fleischfresser, Nagetiere, Fledermäuse; vgl. Abb. 4 über die Bauplantypen des Incudomallear-Gelenkes bei lebenden Säugetieren).
Microtherulum stellt den frühesten bekannten fossilen Beleg für die Existenz eins modernen sattelartigen Hammer-Amboss-Gelenks dar.
Figure 4. Abb. 4 Bauplantypen des Hammer-Amboss-Gelenkes in einer Ansicht von hinten bei (v. l. n. r.): Kloakentieren (Schnabeltier), Beuteltieren und Plazentatieren: vermeintlicher „Urtyp“ (Opossums), „Übergangstyp“ (Hamster), Mikrotyp-Theria-Ohr (Mäuse, Fledermäuse, Microtherulum oneirodes) und frei beweglicher Typ (Eichhörnchen). Bei jedem Bauplantyp zeigt der Punkt das geschätzte Massenzentrum der Gehörknöchelchenkette. (Nach Wang & Wang 2023, CC BY 4.0 Deed, http://creativecommons.org/licenses/by/4.0/)
Die Gelenkverbindung zwischen Hammer und Amboss von Microtherulum stellt den frühesten bekannten fossilen Beleg für die Existenz eines sattelartigen Hammer-Amboss-Gelenks dar. Daraus muss geschlussfolgert werden, dass diese für die modernen Säugetiere charakteristische Gelenkkonfiguration bereits bei ihren ältesten bekannten Vertretern (Eutheriern) in der unteren Kreide ausgebildet war.
Konvergenz und paralleles Auftreten der Mittelohrstrukturen
Wichtig zu erwähnen ist, dass diese moderne Konfiguration des Mittelohres bei Microtherulum oneirodeszeitgleich zu anderen Formvarianten bei den Schwestergruppen der modernen Säugetiere existierte (Haramiyida, Multituberculata, Eutriconodonta, Zhangheotheriidae und Monotremata [Kloakentiere]). Damit ist es in biostratigrafischer Hinsicht nicht mehr möglich, von „primitiveren“ und „modernen“ Varianten der MOK zu sprechen. Ein häufig als primitiv gewertetes Merkmal bei diesen zeitgleich lebenden Schwestergruppen ist ein teilweise verknöcherter Meckelscher Knorpel wie auch eine spezielle Furche im Unterkiefer für den Meckelknorpel – und wird deshalb als morphologisches Bindeglied bei der Mittelohrentwicklung bei Säugetieren dargestellt. Diese Struktur fehlt beim zeitgleich lebenden Microtherulum oneirodesvollständig.Die Autoren Wang & Wang (2023) ziehen daraus den Schluss: „Es scheint, dass der Abbau des Meckelschen Knorpels auch bei den Eutheriern der frühen Kreidezeit variabel ist, was auf eine hohe evolutionäre Variabilität bei der Ablösung des Mittelohrs bei den frühen Säugetierformen mit Ausnahme der Allotherier hinweist.“
Aus evolutionärer Perspektive müssen vielfache, unabhängige und gleichgerichtete Entwicklungssprünge angenommen werden.
Abb. 5 dokumentiert die große Variabilität der Mittelohrformen bei den Säugetieren im weiteren Sinne (Mammaliaformes). Darüber hinaus wird erneut belegt, dass aus evolutionärer Perspektive vielfache, unabhängige und gleichgerichtete Entwicklungssprünge bei der Entstehung des Mittelohres angenommen werden müssen. Das freistehende Mittelohr (detached middle ear, DME) und die damit verbundene Entkopplung von Hör- und Kauapparat entwickelte sich nach der aktuellen Phylogenie mehrfach in mesozoischen Säugetierformen (Abb. 5). Als zugrundeliegender Mechanismus für das Entkopplungsereignis (wobei man hier richtiger von einem hypothetischen Selektionsvorteil sprechen sollte), wird von den Autoren lediglich die Anpassung an ein effizientes Kauen ins Spiel gebracht.
Figure 5. Abb. 5 Hypothetische Evolutionsstufen der Mittelohrregion in einer vereinfachten Phylogenie der Säugetiere inkl. der evolutionären Entwicklung zu den modernen tribosphenischen Backenzähnen (s. unten links: einfache Form: prätribosphenische Gruppe, Übergangsform: vielhöckrige Zahn-Gruppe, säugetiertypische Form: tribosphenische Gruppe), sowie der Abtrennung des Mittelohrs vom Unterkiefer durch die Umwandlung von Unterkieferknochen bei Reptilien (Quatratum, Artikulare, Angulare) zum Hammer-Amboss-Gelenk. Die verschiedenen Elemente des Mittelohrs sind markiert, das Ektotympanon in Lila, der Hammer (Malleus) in Gelb, der Amboss (Incus) in Grün und der verknöcherte Meckel‘sche Knorpel in Orange. Man beachte das konvergente, gleichzeitige und zum Teil rückschrittliche Auftreten der tribosphenischen bzw. der prätribosphenischen Formen, des DME, des MAME, eines verknöcherten Meckel‘schen Knorpels und des überlappenden Gelenkes zwischen Hammer und Amboss in den unterschiedlichen rezenten und ausgestorbenen Säugetiergruppen. (Nach Wang & Wang 2023, CC BY 4.0 Deed, http://creativecommons.org/licenses/by/4.0/).
Diskussion
Der aktuelle Fossilfund des modernen Säugetiers Microtherulum oneirodes aus der Unterkreide bestätigt, dass aus evolutionärer Perspektive davon ausgegangen werden muss, dass auch die Entstehung des Mittelohres und seiner Begleitstrukturen bei modernen Säugetieren und ihren fossilen Verwandten mehrfach und unabhängig voneinander erfolgt sein muss. Außerdem hilft das Fossil nicht aufzuklären, von welchen Vorfahren die für moderne Säugetiere typische Konfiguration der Gelenkverbindungen zwischen Hammer und Amboss (Incudomalleolar-Gelenk) abzuleiten ist, da diese in den frühesten Formen bereits völlig modern erscheint. Die zugrundeliegenden Mechanismen für die postulierten konvergenten Entwicklungen und die dafür erforderlichen Selektionsbedingungen der Mittelohrknochen bei Säugetieren bleiben auch nach dem Fund von Microtherulum oneirodes unbekannt. Der Fund von Microtherulum oneirodes ist ein erneutes Beispiel für das allgegenwärtige und kausal ungeklärte Phänomen der „korrelierenden“/konvergenten Merkmalsevolution in der Säugetierevolution. Die damit verbundene Untauglichkeit von Homologien (Merkmalsähnlichkeiten) bei der Rekonstruktion makroevolutionärer Vorgänge wird erneut exemplarisch bestätigt. Dieser Zusammenhang wiegt umso schwerer, da Homologien nach wie vor als das wesentlichste Argument für die gemeinsame Abstammung der Lebewesen aus dem Blickwinkel der Darwin‘schen Evolution gelten.
Literatur
Bertolini R & Leutert G (1982) Atlas der Anatomie des Menschen Nach systematischen und topographischen Gesichtspunkten. Band 3: Kopf und Hals, Gehirn, Rückenmark und Sinnesorgane. Leipzig.
Luo ZX et al. (2007 a) A new eutriconodont mammal and evolutionary development in early mammals. Nature 446, 288–293.
Luo, ZX. (2007 b) Transformation and diversification in early mammal evolution. Nature 450, 1011–1019.
Luo ZX (2011) Developmental patterns in Mesozoic evolution of mammal ears. Annu. Rev. Ecol., Evol. Syst. 42, 355–380.
Luo ZX, Crompton AW & Sun AL (2001) A new mammaliaform from the early Jurassic and evolution of mammalian characteristics. Science 292, 1535–1540.
Meng J, Wang Y & Li C (2011) Transitional mammalian middle ear from a new Cretaceous Jehol eutriconodont. Nature 472, 181–185.
Springer MS et al. (2004) Molecules consolidate the placental mammal tree. Trends Ecol. Evol. 19, 430–438.
Takechi & Kuratani (2010) History of studies on mammalian middle ear evolution. J. Exp. Zool. 314B, 417–433.
Ullrich H (1994) Embryologie und Homologie – Die Reichert-Gauppsche Theorie. Stud. Int. J. 1, 15–24.
Ullrich H (2009) Das Ende der klassischen Homologisierung. Neue Methoden in der vergleichenden embryonalen Forschung. Stud. Integr. J. 12, 58–65.
Ullrich H (2012) Verwirrende Konvergenzen. Zur Evolution des Mittelohres der Säugetiere. Stud. Integr. J. 19, 20–33.
Ullrich H (2013) Moleküle und Anatomie im Konflikt? Die Systematik der Säugetiere. Stud. Integr. J. 20, 119–121.
Ullrich H (2015) Wann entstanden die modernen Säugetiere? Einsichten aus Fossilien, Molekülen und Datierungen. Stud. Integr. J. 22, 23–30.
Wang H & Wang Y (2023) Middle ear innovation in Early Cretaceous eutherian mammals. Nat. Commun. 14:6831, 1–12.
Wang J. et al. (2021) A monotreme-like auditory apparatus in a Middle Jurassic haramiyidan. Nature 590, 279–283.